
Роль взаимодействия полиморфных локусов гена FLG в формировании хронической истинной экземы у женщин
Aннотация
Актуальность: Экзема является мультифакториальным заболеванием, развитие которого определяется полигенной основой и действием провоцирующих факторов экзогенной и эндогенной природы. Цель исследования: Проанализировать роль взаимодействия полиморфных локусов гена филаггрина (FLG) в формировании хронической истинной экземы у женщин. Материалы и методы: Для исследования была сформирована выборка, включающая 446 женщин, в том числе 233 пациентки с хронической истинной экземой и 213 женщин без данного заболевания. Все женщины, включенные в исследование, прошли необходимое клиническое и клинико-лабораторное обследование. Выполнено генотипирование девяти полиморфных локусов гена филаггрина. SNP×SNP взаимодействия, вовлеченные в формирование хронической истинной экземы, изучались с помощью программного обеспечения АРSampler (использовался метод Монте-Карло марковских цепей и байесовская непараметрическая статистика) и MDR (основан на методе снижения размерности MDR). Результаты: В состав двух- и трех-локусных моделей, ассоциированных с формированием заболевания, входят пять полиморфных локусов – rs61816761, rs4363385, rs77199844, rs471144 и rs558269137 гена FLG. Полиморфизмы rs4363385 и rs77199844 включены в наибольшее количество моделей (по 6 моделей соответственно). SNP×SNP взаимодействия rs471144× rs77199844 и rs471144× rs4363385 определяют -0,84% и -0,77% энтропии признака соответственно. Полиморфные локусы rs77199844 и rs471144 локализованы в регионе регуляторных мотивов ДНК к 12 транскрипционным факторам, а полиморфизм rs4363385 гена FLG ассоциирован с уровнем экспрессии девяти генов в коже (SPRR1B, LCE3C, LCE1D, SPRR2D, SPRR2B, LCE3A, LCE1E, SPRR1A, SPRR3). Заключение: Взаимодействия полиморфных локусов гена FLG ассоциированы с формированием хронической истинной экземы у женщин.
Ключевые слова: полиморфизм, ассоциации, SNP×SNP взаимодействия, хроническая истинная экзема, женщины, FLG
Введение. Экзема – острое или хроническое рецидивирующее аллергическое заболевание кожи, формирующееся под влиянием экзогенных и эндогенных триггерных факторов и характеризующееся появлением полиморфной сыпи, острой воспалительной реакцией, обусловленной серозным воспалением кожи, и сильным зудом [1]. В зарубежных литературных источниках синонимом экземы является атопический дерматит [MIM 603165] [2, 3]. Удельный вес экземы составляет до 30-40% в структуре всех заболеваний кожи [1]. При дерматозах утрата временной трудоспособности достигает до 36%. Среди госпитализированных пациентов дерматологического профиля на долю больных экземой приходится более 30% [4].
Экзема является мультифакториальным заболеванием, развитие которого определяется полигенной основой (наследственная предрасположеность обусловливает различные нарушения со стороны нервной, эндокринной, иммунной систем) и действием провоцирующих факторов экзогенной (бактериальные и грибковые инфекционные агенты, химические вещества, физические факторы, лекарственные препараты, пищевые продукты) и эндогенной (антигены микроорганизмов из очагов хронической инфекции) природы [1, 5]. Наследственные факторы в значительной степени определяют подверженность к развитию экземы [3, 6-8]. Вероятность возникновения заболевания составляет около 40% при наличии экземы у одного из родителей, и этот показатель достигает уровня 50-60% при наличии заболевания у обоих родителей [1]. К настоящему времени по данным GWAS каталога проведено десять полно-геномных исследований атопического дерматита (атопической экземы), в результате которых выявлено более 100 GWAS-значимых полиморфных локусов, вовлеченных в формирование заболевания (https://www.ebi.ac.uk/gwas/efotraits/EFO_0000274). Важный вклад в формирование подверженности к развитию экземы вносят мутации в гене филаггрина [3, 7-11]. Различными научными коллективами активно изучаются ассоциации мутаций гена FLG, связанных с потерей функции (loss-of-function variants – p.Arg501∗(c.1501C>T), p.Ser761Cysfs∗36 (c.2282_2285del), p.Arg2447∗ (c.7339C>T), p.Ser3247∗ (c.9740C>A), R501X, 2282del4, R2447X, S3247X и др.) с формированием экземы (атопического дерматита) [3, 12, 13]. Следует отметить, что в России генетические исследования, посвященные этому вопросу единичные [14, 15].
Цель исследования. Анализ роли взаимодействия полиморфных локусов гена FLG в формировании хронической истинной экземы у женщин.
Материалы и методы исследования. Для исследования была сформирована выборка, включающая 446 женщин, в том числе 233 пациентки с хронической истинной экземой и 213 женщин без данного заболевания (контрольная группа). Все индивидуумы, включенные в исследование, родились и проживали в Центральном Черноземье России (Курская, Белгородская, Воронежская, области) [16], имели русскую национальность и не являлись родственниками. Из исследования были исключены пациенты с другими формами экземы; больные, у которых хроническая истинная экзема сочеталась с иной кожной патологией или соматическим заболеванием, влияющим на состояние кожи; пациенты, имеющие тяжелую соматическую патологию (онкологические заболевания, ревматоидный артрит, болезнь Крона), а также пациенты, регулярно принимающие антигистаминные, стероидные, иммуносупрессивные препараты; индивидуумы моложе 18-летнего возраста; индивидуумы, отказавшиеся от проводимого исследования.
Диагноз ХИЭ устанавливался на основании жалоб, анамнеза, клинических проявлений, течения заболевания и лабораторных методов исследования. Контрольная группа формировалась из индивидуумов, не имеющих заболеваний кожи на момент обследования, а также соматической патологии, приводящей к вторичному поражению кожи. Клиническое и клинико-лабораторное обследование больных проводилось на базе поликлинического отделения ОБУЗ "Курский областной клинический кожно-венерологический диспансер". Выборки формировались за период с 2010 по 2016гг. Средний возраст больных хронической истинной экземой составил 46,71 ± 2,46 (варьировал от 18 лет до 83 лет), контрольной выборки- 44,68 ±2,76 (варьировал от 20 до 72 лет) (р>0,05). Исследование проводилось с информированного согласия индивидуумов на его проведение.
В исследуемых группах женщин (с ПЭ и контрольная группа) проведено молекулярно-генетическое исследование десяти полиморфных локусов гена филаггрина – rs471144, rs61816761, rs4363385, rs558269137, rs77199844, rs12144049, rs3126085, rs10888499, rs6661961. Полиморфные локусы включались в исследование в соответствии с их ассоциацией с экземой (атопическим дерматитом) по данным ранее проведенных полногеномных (GWAS) исследований, значимым регуляторным потенциалом и влиянием на экспрессию генов. Генотипирование проводилось на амплификаторе CFX-96 Real-Time System (Bio-Rad) с помощью полимеразной цепной реакции (ПЦР) синтеза ДНК в режиме реального времени методом TаqMаn зондов с использованием наборов реагентов для амплификации ДНК с соответствующими олигонуклеотидными праймерами и зондами для изучаемых полиморфных локусов гена филаггрина, синтезированных ООО «ТестГен» (Ульяновск).
В настоящем исследовании изучена роль взаимодействия полиморфных локусов гена филаггрина в возникновении ХИЭ у женщин. SNP×SNP взаимодействия, вовлеченные в формирование ХИЭ, изучались с помощью двух подходов. Во-первых, использовали программное обеспечение АРSampler (http://sources.redhat.com/cygwin/), использующий метод Монте-Карло марковских цепей и байесовскую непараметрическую статистику [17, 18]. Силу ассоциации оценивали показателем отношения шансов (OR) с 95% доверительным интервалом (95%CI). Выполнен пермутационный тест с целью коррекции на множественные сравнения - статистически значимым уровнем являлся pperm<0,05. Во-вторых, использовали метод снижения размерности MDR (Multifactor Dimensionality Reduction) (http://www.multifactordimensionalityreduction.org/). Расчеты проводились в программе MDR (v. 3.0.2) (http://sourceforge.net/projects/mdr).
Регуляторный потенциал исследуемых полиморфных локусов гена FLG изучался с помощью программы HaploReg (v4.1) (http://archive.broadinstitute.org/mammals/haploreg/haploreg.php) согласно методике указанной в работе [19].
Рассмотрена связь полиморфизма гена FLG с экспрессией генов (cis-eQTL) по данным проекта Genotype-Tissue Expression (GTEx) (http://www.gtexportal.org/). В работу включались данные с p<8*10-5, pFDR≤0,05. Оценка связи аллельных вариантов рассматриваемых полиморфных локусов с уровнем транскрипции генов проводилось по методике, представленной в работе [20].
Результаты и их обсуждение. В таблице 1 представлены полученные данные о распределение генотипов в исследуемых выборках женщин (больные и контроль), наблюдаемой (Ho) и ожидаемой гетерозиготности (He), соответствии наблюдаемого распределения генотипов ожидаемому при равновесии Харди-Вайнберга (РHWE) по изучаемым локусам гена филаггрина

С помощью программного обеспечения АРSampler установлены сочетания аллелей/генотипов полиморфных локусов генов филаггрина, ассоциированные с развитием хронической истинной экземы у женщин (таблица 2). Среди изученных полиморфных локусов гена филаггрина в состав моделей SNP×SNP взаимодействий, связанных с формированием заболевания, входят пять полиморфных локусов – rs61816761, rs4363385, rs77199844, rs471144 и rs558269137. Нами выявлено 5 двухлокусных и 4 трехлокусных моделей SNP×SNP взаимодействий, определяющих подверженность к развитию ХИЭ. При этом следует отметить, что, во-первых, двух-локусная модель, включающая полиморфные локусы rs471144 × rs61816761 имеет наибольшую статистическую значимость (p=0,003, pperm=0,0001). Во-вторых, среди 9 статистически значимых 2-х и 3-х локусных моделей, ассоциированных развитием ХИЭ, 8 моделей имеет протективное значение и лишь одна модель (аллель G rs471144 × аллель G rs61816761) является фактором риска развития заболевания (OR=6,66, 95%CI 1,14-38,91). В третьих, в наибольшее количество моделей, определяющих подверженность к развитию ХИЭ, входят полиморфные локусы rs4363385 и rs77199844 (по 6 моделей соответственно)

С использованием метода Multifactor Dimensionality Reduction нами установлена статистически значимые 4-х и 5-и локусные модели SNP×SNP взаимодействий гена филаггрина, ассоциированные с развитием хронической истинной экземы. Четырех-локусная модель включает полиморфизмы rs471144, rs4363385, rs558269137, rs77199844. Показатель Training Bal. Acc. данной модели составил 55,08%, Testing Bal. Acc. - 45,25%, воспроизводимость модели CVC=8/10, OR=1,63, 95%CI 1,07-2,50 (рperm<0,001), чувствительность модели – 0,31, специфичность модели – 0,78. В пяти-локусную модель входят полиморфные локусы rs471144, rs61816761, rs4363385, rs558269137, rs77199844. Показатели точности данной модели составили Training Bal. Acc. - 55,47%, Testing Bal. Acc. - 48,39%, воспроизводимость CVC=10/10, OR=1,70, 95%CI 1,11-2,59 (рperm<0,001), чувствительность – 0,32, специфичность – 0,78. Дендрограмма и граф SNP×SNP взаимодействий наиболее лучшей пяти-локусной модели гена филаггрина представлены на рисунке 1. Следует отметить, что, во-первых, наиболее значимые SNP×SNP взаимодействия (отличаются наибольшими показателями энтропии) характерны для полиморфных локусов rs471144× rs77199844 (данное взаимодействие определяет -0,84% энтропии) и rs471144× rs4363385 (определяет -0,77% энтропии). Во-вторых, для SNP×SNP взаимодействий rs471144×rs77199844 и rs471144×rs4363385 характерны выраженные антагонистические взаимодействия (на дендрограмме и графе линии их соединяющие окрашены в синий цвет).

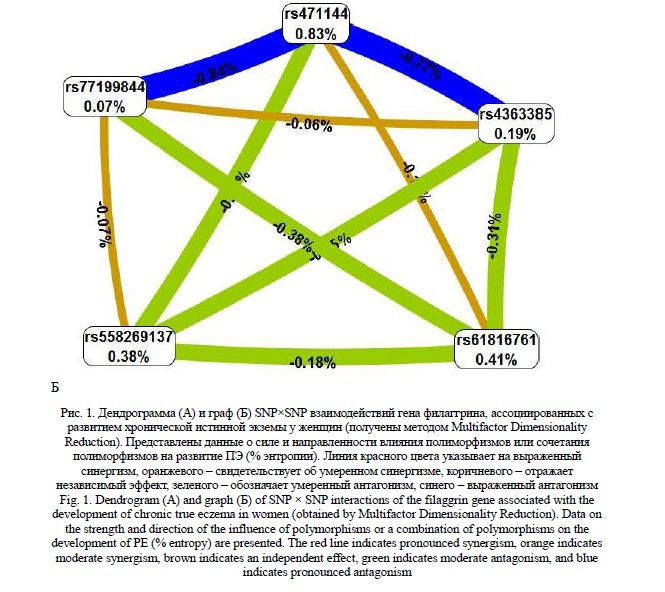
Анализ регуляторного потенциала «наиболее значимых» для развития ХИЭ полиморфных локусов, проведенный с помощью программы HaploReg (v4.1) показал, что полиморфизм rs77199844 гена FLG находится в регионе модифицированных гистоновых белков (H3K4me1), маркирующих энхансеры в культуре клеток «ES-I3 Cells» и первичных клетках дермальных фибробластов (NHDF-Ad Adult Dermal Fibroblast Primary Cells), а также расположен в регионе регуляторных мотивов ДНК, определяющих взаимодействие с семью транскрипционными факторам (Foxa, Foxd1, Foxf1, Foxf2, Foxq1, HDAC2, p300). Следует отметить, что альтернативный аллель С rs77199844 снижает афинность ко всем этим семи транскрипционным факторам (различия между LOD коэффициентами альтернативного и референсного аллелей отрицательные). Наряду с этим полиморфный локус rs471144 так же локализован в области модифицированных гистоновых белков (H3K4me1, H3K27ac), маркирующих энхансеры в первичных клетках Т-хелперов в периферической крови и регионе мотивов ДНК, влияющих на афинность к 7 факторам транскрипции (Fox, Foxo, Foxj2, Sox, Foxf2, Zec, SREBP). К подавляющему большинству этих транскрипционных факторов (за исключением TF – SREBP) альтернативный аллель G снижает афинность. Таким образом, два полиморфных локуса rs77199844 и rs471144, вовлеченные в формирование ХИЭ, локализованы в регионе регуляторных мотивов ДНК к 12 транскрипционным факторам, что свидетельствует о их важном регуляторном потенциале.
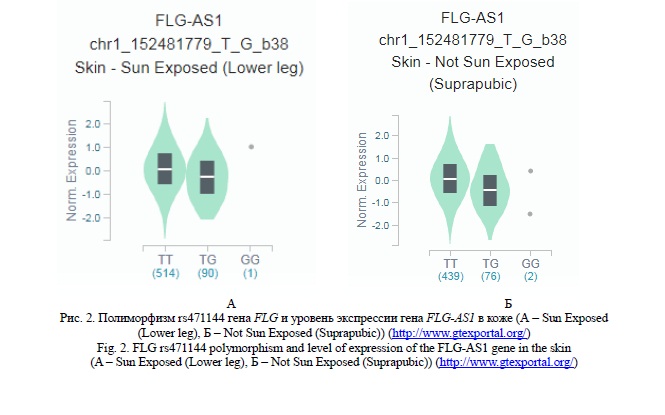
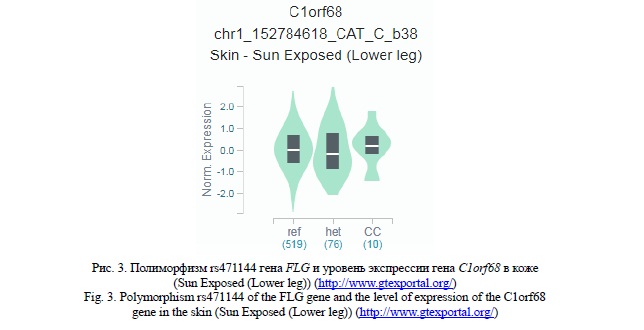
С использование данных проекта Genotype-Tissue Expression (GTEx) (http://www.gtexportal.org/) установлено важное eQTL значение полиморфных локусов rs471144, rs77199844 и rs4363385, имеющих наибольший вклад в подверженность к развитию ХИЭ. Выявлены ассоциации полиморфизма rs471144 с уровнем транскрипционной активности гена FLG-AS1 в коже (рисунок 2). При этом, следует отметить, что аллель G связан с пониженной экспрессией гена FLG-AS1 (β= -0,34 – -0,44, р≤3,5*10-5, pFDR≤0,05). Полиморфный локус rs77199844 ассоциирован с уровнем экспрессии гена C1orf68 в коже (рисунок 3) - аллель С определяет более низкую экспрессией данного гена (β=-0,16, р=2,9*10-5, pFDR≤0,05). Наибольшее eQTL значение имеет полиморфизм rs4363385 гена FLG (таблица 3). Данный полиморфный локус ассоциирован с уровнем экспрессии девяти генов в коже (SPRR1B, LCE3C, LCE1D, SPRR2D, SPRR2B, LCE3A, LCE1E, SPRR1A, SPRR3). Обращает на себя внимание факт того, что полиморфный вариант С полиморфизма rs4363385 гена FLG определяет повышенную экспрессию генов SPRR2D, SPRR1A, SPRR3,LCE3A (β<0, pFDR≤0,05 ) и низкую экспрессию генов SPRR1B, LCE3C, LCE1D, SPRR2B и LCE1E (β >0, pFDR≤0,05 ) (таблица 3).

Ген филаггрина состоит из трех экзонов и двух интронов и расположен в кластере генов комплекса эпидермальной дифференцировки (1q21) [21]. Филаггрин образуется из профилаггрина и является ключевым белком, способствующим терминальной дифференцировке эпидермиса и образованию защитного барьера кожи. Профилаггрин кодируется третьим экзоном [22]. Установлено, что мутации, приводящие к потере функции филаггрина (loss-of-function variants) или нулевые мутации гена филаггрина, обусловливают образование неактивной формы синтезированного полипептида (вследствие преждевременного прекращения синтеза, сигналом для которого являются появившиеся в результате мутаций стоп-кодоны). Это в конечном итоге приводит к низкой концентрации профилаггрина в зернистом слое эпидермиса, что в последующем определяет формирование аномально тонкого слоя кератиноцитов, и является морфологическим субстратом, лежащим в основе предрасположенности к хроническим заболеваниям кожи (дерматит, экзема, псориаз и др) [23-25]. В нашей работе выявлены ассоциации с развитием хронической истинной экземы у женщин как мутаций, связанных с потерей функции филаггрина (2282del4 - rs558269137, R501X - rs61816761) так и полиморфных локусов гена филаггрина (rs4363385, rs77199844 и rs471144). Значимые ассоциации с развитием экземы (атопического дерматита) этих мутаций и полиморфных локусов показаны в ранее проведенных работах [3, 5-11, 14-15]. Так, в результате полно-геномного исследования атопического дерматита и псориаза, проведенного в 2015г Baurecht H. et al. [3] установлены ассоциации с риском развития атопического дерматита rs77199844 (del) (OR=2,01 95%CI 1,72–2,35), rs4363385 (OR=1,23 95%CI 1,15–1,32) и rs471144 (OR=1,54 95%CI 1,37–1,73). Так же в этой работе показана вовлеченность в формирование псориаза на полно-геномном уровне полиморфных локусов rs77199844 (del) (OR=1,16 95%CI 1,01–1,33) и rs4363385 (OR=0,89 95%CI 0,85–0,94). Полиморфизм rs471144 не был ассоциирован с псориазом (OR=1,03 95%CI 0,94–1,14). Авторы этого исследования обращают внимание на факт «обратной» ассоциации полиморфизма rs4363385 с риском развития атопического дерматита и псориаза: для атопического дерматита этот полиморфизм является фактором риска (OR=1,23), а для псориаза – наоборот, протективным фактором (OR=0,89). Следует отметить, что ряд из исследуемых нами мутаций гена филаггрина (например, R501X (rs61816761)) наряду с вовлеченностью в подверженность к заболеваниям кожи (дерматит, экзема) показывают ассоциации на полно-геномном уровнем и с другими заболеваниями со значимой иммуно-аллергической компонентой (бронхиальная астма) [26, 27].
Заключение. Взаимодействия полиморфных локусов гена FLG ассоциированы с формированием хронической истинной экземы у женщин. В состав двух- и трех-локусных моделей, определяющих подверженность к заболеванию, входят пять полиморфных локусов - rs61816761, rs4363385, rs77199844, rs471144 и rs558269137 гена FLG. Полиморфизмы rs4363385 и rs77199844 включены в наибольшее количество моделей (по 6 моделей соответственно). SNP×SNP взаимодействия rs471144× rs77199844 и rs471144× rs4363385 определяют -0,84% и -0,77% энтропии признака соответственно. Полиморфные локусы rs77199844 и rs471144 локализованы в регионе регуляторных мотивов ДНК к 12 транскрипционным факторам, а полиморфизм rs4363385 гена FLG ассоциирован с уровнем экспрессии девяти генов в коже (SPRR1B, LCE3C, LCE1D, SPRR2D, SPRR2B, LCE3A, LCE1E, SPRR1A, SPRR3).
В отношении данной статьи не было зарегистрировано конфликта интересов.













Список литературы