
HSPD1 gene polymorphism is associated with an increased risk of ischemic stroke in smokers
Aннотация
Background: Ischemic stroke (IS) is a leading cause of death and disability worldwide. Since the influence of vascular stressors prompts cross-reactivity with microbial and human HSPD1 (HSP60), which results in the degradation of vascular endothelium and intimal infiltration by mononuclear cells, HSP60 is viewed as a key player in the autoimmune theory of atherosclerosis. The aim of the study:The present pilot study investigated whether the tag SNP rs11682567 HSPD1 is associated with the risk of IS. Materials and methods: DNA samples from 1176 unrelated Russian subjects (708 IS patients and 468 healthy controls) were genotyped for tagging SNP rs11682567 in the HSPD1 gene using the MassArray-4 system. A log-additive regression model was used to interpret the associations between genotypes and IS risk. Results: We discovered that smoking, a major environmental endothelial stressor, dramatically modifies the link between rs11682567 and IS, with a relationship only being seen in smokers (risk allele G; OR = 2.11, 95% CI = 1.17-3.82; P = 0.0075). The functional annotation of this SNP revealed that risk allele G of rs11682567 impacts the expression and alternative splicing of genes that regulate brain ischemia-reperfusion injury, reactive oxygen species production, toll-like receptor 4 internalization, and pro-inflammatory properties. Moreover, allele G rs11682567 creates binding sites for transcription factors jointly involved in biological processes encompass neurogenesis, monocyte differentiation, fatty acid metabolism, cell signalling, and apoptosis. Conclusion: Thus, rs11682567 SNP in the HSPD1 gene represents a novel genetic marker of ischemic stroke, whose significance for the pathogenesis of IS is mediated by smoking.
Ключевые слова: ischemic stroke, heat shock, mitochondrial folding, HSPD1, HSP60, chaperones, atherosclerosis, autoimmune response, rs11682567, smoking
К сожалению, текст статьи доступен только на Английском
Introduction. Ischemic stroke is a major global cause of death and disability [1]. A lot of studies have been done so far to investigate the genetic nature of stroke [2] and stroke-related phenotypes, like hypertension [3] and atherosclerosis [4]. Heat Shock Proteins (HSPs) are engaged in a number of cellular processes, including immunological response, inflammation, and apoptosis, that may have an effect on the outcomes of stroke [5]. Also, by stabilising proteins and assisting in protein refolding and degradation, HSPs directly contribute to the protection of cells and tissues against the damaging effects of ischemia [6].
HSPD1, otherwise known as heat shock protein 60 (HSP60), is a chaperonin that is present in human mitochondria. It assists in protein folding and is required for preserving mitochondrial functionality, which is essential for cell survival [7]. A major risk factor for stroke is atherosclerosis, an inflammatory condition which was initially referred to an immunological response against HSP60 [8]. There is a chance of cross-reactivity between human and microbial HSP60, which can cause endothelial damage and inflammation and possibly aid in the onset of atherosclerosis [4]. To our knowledge, no research has looked into the relationship between HSPD1 SNPs and IS risk.
The aim of the study. To investigate whether SNP rs11682567 HSPD1 is related with the risk of IS.
Materials and methods. The study enrolled 1176 unrelated Russians from Central Russia, including 708 patients with IS and 468 healthy people. The study protocol was approved by the Kursk State Medical University's Ethical Review Committee. Before being accepted into the study, each participant provided written informed consent and agreed to the inclusion criteria listed below: self-declared Russian ancestry and birthplace in Central Russia.
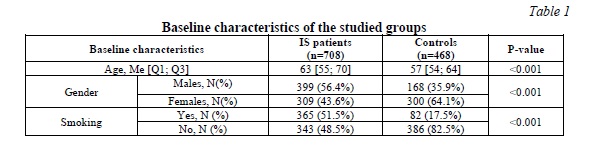
In Table 1, the study population's baseline characteristics are presented.
Both the Regional Vascular Centre of Kursk Regional Clinical Hospital and the Neurology Clinics of Kursk Emergency Medicine Hospital participated in the study's patient recruitment [9-12]. In order to diagnose IS during the acute phase of stroke, the results of the neurological examination, brain computed tomography, and/or magnetic resonance imaging were used. The following conditions were used to exclude individuals from the IS study: traumatic brain injury; intracerebral haemorrhage; hemodynamic or dissection-related stroke; hepatic or renal failure; autoimmune, oncological, or other disorders that can produce an abrupt cerebrovascular event. All of the IS patients had a history of hypertension and were taking medication to lower their blood pressure.
The control group consisted of healthy volunteers with normal blood pressure, not receiving antihypertensive therapy, without clinical signs of cardiovascular, cerebrovascular and other serious diseases. If a healthy individual's systolic blood pressure was less than 130 mm Hg and/or their diastolic blood pressure was less than 85 mm Hg on at least three different occasions, they would have qualified for inclusion in the control group. When undergoing medical examinations at public institutions and industrial businesses in the Kursk region, control subjects were recruited from Kursk hospitals during particular periods [13, 14]. During the same time frame and from the same population, this cohort was recruited [15].
The selection of SNPs was based on the following criteria: the SNP needs to be tagging, with a minor allele frequency of at least 0.05 in the European population, and stand out for having a high regulatory potential. The bioinformatic tools SNPinfo Web Server (https://snpinfo.niehs.nih.gov/) and LD TAG SNP Selection, which were used to identify SNPs based on the reference haplotypic structure of the Caucasian populations (CEU, TSI) of the project HapMap, determined that the rs11682567 HSPD1 is a tagging SNP. This SNP is located in intron chr2:197490429 (GRCh38.p14) and characterized by the replacement T>G (https://www.ncbi.nlm.nih.gov/snp/rs11682567).
The regulatory potential of rs11682567 was evaluated using a variety of bioinformatic tools [16]. The RegulomeDB online tool (https://regulomedb.org/regulome-search/ (accessed November 20, 2022)) showed that rs11682567 is characterized by a regulatory coefficient of 5 (TF binding or DNase peak). This SNP is characterized by Enhancer histone marks and regulatory motifs, according to the HaploReg (v4.1) database (http://archive.broadinstitute.org/mammals/haploreg/haploreg.php (accessed on November 20, 2022)).
The NCBI source's data (https://www.ncbi.nlm.nih.gov/snp/) indicates that this genetic variation is described by an average minor allele frequency of >0.05 in European populations. In light of the fact that SNP rs11682567 satisfies the prerequisites for study inclusion, it was chosen for our research.
Genetic Analysis. The Laboratory of Genomic Research at the Research Institute for Genetic and Molecular Epidemiology of Kursk State Medical University (Kursk, Russia) performed the DNA analysis. The conventional technique of phenol/chloroform extraction and ethanol precipitation was used to extract genomic DNA from thawed blood samples. The MassARRAY-4 genetic analyzer (Agena Bioscience, San Diego, CA, USA) was used for genotyping.
The SNP primers were created through the use of the Agena Bioscience's Assay Design Suite software, and genotyping was accomplished by means of matrix-assisted laser desorption ionization mass spectrometry (MALDI-TOF). The remaining dNTP and primers were removed by employing the SAP enzyme after a PCR reaction specific to the locus. Mass-modified dideoxy-nucleotide terminators of an oligonucleotide primer were then utilized to carry out single-base extension, which occurred upstream of the desired polymorphic site.
A PCR reaction was conducted on a spectral chip using a 5 µL reaction mixture that included 3 µL of PCR mix consisting of 80.6 µL of ddH2O, 50.4 µL of 10x buffer, 40.3 µL of MgCl2 (25 mmol/L), 10.1 µL of a mixture of dNTP (25 mmol/L), 100.8 µL of a mixture of PCR primers (0.5 mmol/L), and 20.16 µL of HotStartTaq polymerase (5 U/µL), as well as 2 µL of DNA (10 ng/µL). The reaction was carried out for 125 minutes in a CFX96 Bio-Rad thermal cycler, with the following program: denaturation at 95 °C for 2 minutes, followed by 44 cycles of denaturation at 95 °C for 30 seconds, annealing at 56 °C for 30 seconds, and elongation at 72 °C for 60 seconds. The samples were then incubated at 72 °C for 5 minutes.
The unreacted dNTPs present in the PCR samples were eliminated using the shrimp alkaline phosphatase (SAP) reaction, which involved the hydrolysis of dNTPs to nucleosides and phosphates by the alkaline phosphatase of shrimp. To achieve this, 2 µL of SAP mixture, consisting of 161.6 µL of ddH2O, 18 µL of 10xTS buffer, and 31.7 µL of SAP enzyme, was added to each well, and the plate was loaded into the CFX96 Bio-Rad thermal cycler. The SAP reaction was carried out for 55 minutes, comprising 45 minutes at 37 °C, followed by 10 minutes at 85 °C.
To prepare for the third iPLEX reaction, an E-primer mix was created based on the molecular weight of the E-primers. The resulting mixture was evaluated using mass spectrometry to determine the intensity of the E-primer peaks, and the necessary quantities of E-primers were added to the mix to achieve the same signal intensity. 2 µL of iPLEX-mix (consisting of 65.4 µL of ddH2O, 21.2 µL of iPLEX buffer, 21.2 µL of terminating mixture, 99.3 µL of E-primer mix, and 4.33 µL of iPLEX enzyme) was then added to each well. The plate was loaded into a CFX96 Bio-Rad thermal cycler and underwent denaturation at 94 °C for 30 seconds, followed by 40 cycles of denaturation at 94 °C for 5 seconds, annealing at 52 °C for 5 seconds, and elongation at 80 °C for 5 seconds. The reaction was then incubated at 72 °C for 3 minutes. After the iPLEX reaction was complete, 30 µL of water was added to all wells, and the plate was loaded into a mass spectrometer.
To prevent the formation of contaminant peaks resulting from the binding of small polar molecules and positively charged sodium and potassium cations to oligonucleotides, the amplicons underwent robotic desalination using SpectroCLEAN resin (Agena Bioscience).
The samples were loaded onto a spectral chip with an ionization matrix using the Nanodispenser workstation in automatic mode. The samples formed crystals on the SpectroCHIP, which were then ionized by a high-energy laser, resulting in the ionization of DNA molecules. Subsequently, the ionized DNA samples were passed through a time-of-flight tube with an electric field. By detecting the time-of-flight of analytes, we could determine the genotype of the analytes. Finally, the results of mass spectrometric analysis for each SNP were reviewed using the TyperAnalyzer program from Agena Bioscience.
To ensure quality control, 95 randomly selected DNA samples were chosen for repeat genotyping on the same platform, without knowledge of the case-control status. The repeatability test revealed a 100% concordance rate.
• QTLbase Bioinformatic tool (http://www.mulinlab.org/qtlbase/index.html (accessed February 24, 2023)) was used for analysis of expression quantitative trait loci (eQTLs) in brain, whole blood, and blood vessels. Additionally, for QTL analysis of the peripheral blood effect the eQTL online tool was used.
• STRING database's bioinformatic tools were utilised to analyse the main functional partners of HSPD1 (https://string-db.org/ (accessed February 24, 2023)). The STRING database was also used for analysis of biological processes and molecular functions reflecting interactions HSPD1 with key functionally relevant proteins (accessed February 21, 2023).
• The effect of rs11682567 HSPD1 on the binding of transcription factors (TFs) to DNA based on the presence of the reference/alternative alleles was assessed using the atSNP Function Prediction online tool (http://atsnp.biostat.wisc.edu/search (accessed February 21, 2023)). In accordance with a positional weight matrix-based calculation of the impact of SNPs on how well TFs interact with DNA, certain TFs were added.
• It was possible to assess the joint participation of TFs linked to the reference/SNP alleles in overrepresented biological processes directly associated with the pathogenesis of IS using the Gene Ontology web tool (http://geneontology.org (accessed February 24, 2023)).
• The Comparative Toxicogenomics Database (CTD) resource (http://ctdbase.org (accessed February 24, 2023)) was used for interpretation on environment-associated correlates of HSPD1. Based on information from globally published studies, CTD enables the analysis of particular interactions between genes and chemicals in vertebrates and invertebrates. One chemical and one gene or protein were the subjects of this tool's analysis of binary interactions.
• For a bioinformatic examination of the connections between rs11682567 HSPD1 and stroke-related phenotypes, intermediate phenotypes (such as blood pressure, heart rate etc.), the Cerebrovascular Disease Knowledge Portal (CDKP) (https://cd.hugeamp.org/ (accessed February 24, 2023)) and Cardiovascular Disease Knowledge Portal (https://cvd.hugeamp.org/ (accessed February 24, 2023)) were used.
Results and discussion
Associations of rs11682567 HSPD1 with the Risk of Ischemic Stroke. Table 2 displays the genotype frequencies of rs11682567 HSPD1 in the study groups.
In each case and control groups, the genotype frequencies of all examined SNPs were distributed according to the Hardy-Weinberg equilibrium (P>0.05). The analysis of the total sample did not reveal associations between rs11682567 HSPD1 and IS risk (Table 2).

Subsequent sub-group analysis showed that rs11682567 was associated with an increased risk of IS exclusively in smokers (risk allele G; OR = 2.11; 95% CI = 1.17–3.82; P = 0.0075).
Bioinformatics Analysis. The HSPD1 gene is highly expressed in brain tissues, blood vessels, and whole blood. In brain tissues, gene expression levels of HSPD1 (MeTPM) varies from 77.57 to 159.3; in blood vessels – from 122.4 to 131.0; in whole blood MeTPM=30.69.
QTL-effects. According to the QTLbase, the G allele rs11682567 is associated with eQTL-mediated up-regulation of HSPD1 expression in the brain-hippocampus (beta=0.174; FDR=0.002), as well as up-regulation of ANKRD44 in Blood-T cell CD4+naive (beta=0.02; FDR=0.03) and ANKRD44-IT1 in blood-T cells CD4+ naive (beta=0.038; FDR=0.02)/blood-neutrophils CD16+ (beta=0.015; FDR=0.036) (http://www.mulinlab.org/qtlbase).
Furthermore, using the eQTLGen resource, it was discovered that the G allele rs11682567 had an impact on the expression of the genes AC013264.2, MARS2, RFTN2, COQ10B, SF3B1, and HSPE1 as well as an increase in the expression of the PLCL1 gene in peripheral blood (Table 3).

Using the bioinformatics tool FIVEx, it was feasible to determine how the G rs11682567 allele affects the alternative splicing of the HSPD1, ANKRD44, COQ10B, HSPE1, MOB4, RFTN2, and SF3B1 genes in blood cells and brain tissues by cis-sQTL-mediated effects (https://fivex.sph.umich.edu).
Analysis of transcription factors. Risk allele G of rs11682567 creates binding sites for transcription factors BRCA1, COMP1, FOXO3, FOXO4, HMGA1, IRF7, MYC, RUNX2, SOX10, SOX17, SOX2, Sox6, SOX9, SRY. According to the Gene Ontology resource, these transcription factors are jointly involved in overrepresented biological processes, such as «glial cell fate commitment» (GO:0021781; FDR=0.005); «regulation of monocyte differentiation» (GO:0045655; FDR=0.01); «neuronal stem cell population maintenance» (GO:0097150; FDR=0.014); «negative regulation of fatty acid metabolic process» (GO:0045922; FDR=0.03); «negative regulation of neuron differentiation» (GO:0045665; FDR=0.003); «oligodendrocyte differentiation» (GO:0048709; FDR=0.003); «cellular response to growth factor stimulus» (GO:0071363; FDR=0.02); «cellular response to cytokine stimulus» (GO:0071345; FDR=0.009); «negative regulation of apoptotic process» (GO:0043066; FDR=0.02). The etiopathogenesis of IS is greatly influenced by all of these biological processes. The fact that TFs that bind to the G allele also control «DNA methylation» (GO:0044030; FDR=0.016) and «chromatin organization» (GO:0006325; FDR=0.004) adds to the notion that rs11682567 HSPD1 plays a substantial role in the mechanisms of epigenetic regulation. It is remarkable that no overrepresented GO was discovered when TFs that bind to the reference allele T rs11682567 were examined.
Bioinformatic analysis of the associations of rs11682567 HSPD1 with cerebral stroke and intermediate phenotypes. According to the bioinformatic resources Cerebrovascular Disease Knowledge Portal (CDKP) and Cardiovascular Disease Knowledge Portal (CVDKP), which integrate and evaluate the genetic association data from the greatest consortiums for the research of cardiovascular and cerebrovascular disorders, the risk allele G of rs11682567 HSPD1 is associated with increasing in diastolic blood pressure (beta=0.007; P=0.0007), total cholesterol (beta=0.0033; P=0.033), coronary artery disease in subjects without diabetes (OR=1.09; P=0.043).
Protein-Protein Interactions. Using the STRING database, 10 proteins (proteins of the first shell of interactors) distinguished by the most pronounced interactions with HSPD1 were discovered: HSPE1, HSPA9, TLR4, TLR2, HSP90AA1, HSP90AB1, TST, GRPEL1, TRAP1, DHFR (PPI enrichment p-value=7.46×10-5) (Fig. 1).

It is interesting that the majority of HSPD1's primary functional partners exhibit chaperone and co-chaperone activities, which are strongly associated with proteostasis (by regulation of protein import, misfolding and refolding of unfolded proteins under stress), inflammatory response (by NF-kappa-B activation), folate metabolism (Table 4).

HSPD1 and its primary functional partners jointly participate in biological processes which reflects biological processes crucial to the pathophysiology of IS, such as proteostasis, oxidative stress, inflammation, regulation of extracellular matrix, endothelial dysfunction, heat stress, apoptosis, neurogenesis, sell signaling (https://string-db.org/cgi/network?taskId=bACSg8aUeF2I&sessionId=bjoEZTIHrwUn).
The current study demonstrates for the first time that polymorphism rs11682567 in the gene encoding Heat Shock Protein Family D (Hsp60) Member 1 (HSPD1) is linked to the risk IS and is significantly modified by smoking. HSPD1 is expressed in vessels, brain tissues and blood. This chaperonin is responsible for regulating mitochondrial protein homeostasis and mitochondrial function: it encourages unfolded polypeptides to fold normally in stressful conditions, avoids misfolding, and facilitates imported proteins fold properly [17]. Previous studies have demonstrated a substantial function for HSPD1 in the emergence of cardiovascular diseases. Patients with CVD showed elevated HSP60 levels [18]. Currently, HSP60 is seen as a major player in the autoimmune theory of atherosclerosis [8, 19]. The highly conserved microbial HSP60 antigen is a target of protective, advantageous adaptive immunity that all humans develop [20]. However, exposure to endothelial stressors, such as components of cigarette smoke, causes HSP60 expression in vascular endothelial cells that do not typically express HSP60, which causes a (cross)reaction and destruction of these target cells by preexisting cellular and humoral immunity against HSP60, resulting in intimal infiltration by mononuclear cells [19]. HSP60 also has a sizable impact on the regulation of vascular smooth muscle cells because it activates the IKK/NF-B, ERK/MAPK, and TLR4 pathways, which promote vascular smooth muscle cell migration, thicken the intima, and are important in vascular wall remodeling [18, 21, 22, 23]. Thus, in total, exposure to vascular stressors triggers a "vicious circle" of an autoimmune response leading to the progression of atherosclerosis.
According to bioinformatics analysis utilising the STRING database, the key functional partners of HSPD1 are involved in a number of overrepresented biological processes associated with atherosclerosis, managing not just proteostasis but also oxidative stress, MyD88-dependent toll-like receptor signaling pathway, I-kappaB phosphorylation, production of pro-inflammatory cytokines, nitric-oxide synthase level, macrophage activation and proliferation, extracellular matrix organization, apoptosis, tumor necrosis factor production, neurogenesis. This provides more proof that HSPD1 plays a crucial part in the pathogenetic processes that specifically cause stroke.
We used bioinformatics analysis to support the discovered link because no analysis of correlations of rs11682567 HSPD1 with the development of IS has been conducted so far. First, a large number of genes are impacted by the risk allele G of rs11682567 HSPD1's effects on their level of expression and alternative splicing, including: ANKRD44, which can regulate NF-κB, an important pro-inflammatory signaling pathway [24]; PLCL1, which modulates VEGFR2 signalling to promote angiogenesis in the central nervous system and vascular endothelial cell inflammation [25]; MARS2, regulating ROS production [26]; RFTN2, driving toll-like receptor 4 internalisation [27]; COQ10B, regulating mitochondrial respiratory chain assembly [28], brain ischemia-reperfusion [29] and ROS level [30]; SF3B1, a major regulator of hypoxia [31]; HSPE1, a component in ischemia damage control [32]. Second, it was discovered that the risk allele G of rs11682567 creates binding sites for transcription factors that jointly regulate biological processes with a high pathogenetic significance for IS. These biological processes encompass neurogenesis, monocyte differentiation, fatty acid metabolism, cell signalling (response to growth factor stimulus, to cytokine stimulus), and apoptosis. Third, it was discovered that the risk allele G is associated with an increase in diastolic blood pressure, total cholesterol levels, which are modifiable risk factors for the IS, as well as coronary heart disease, which is pathogenetically related to IS.
Numerous studies have shown that smoking can significantly modify the association of genetic markers with the risk of multifactorial diseases [33], including stroke [9-12]. The effect of cigarette smoke on the up-regulation of HSP60 in humans has been demonstrated in a large number of studies [34, 35]. This discovery further justifies the significance of endothelial stressors in initiating HSPD1-dependent pro-atherosclerotic molecular pathways and helps to explain the strong risk effects of rs11682567 HSPD1 we observed in smokers and their absence in nonsmokers.
In conclusion, our case-control study findings indicate that the HSPD1 gene variant of rs11682567 contributes to IS susceptibility. Further research is required to confirm these findings and look into the potential molecular mechanisms underlying the associations between HSPD1 gene variants and IS risk.
Conclusion. The present study reveals the novel genetic association between rs11682567 HSPD1 and the risk of IS observed exclusively in smokers. These data provide the new insights of involvement of HSP60 in cardiovascular pathology. Further studies may help to uncover the details of its particular role in the stroke pathogenesis.
Financial support
This research was funded by the Russian Science Foundation (№ 22-15-00288, https://rscf.ru/en/project/22-15-00288/).













Список литературы
Список использованной литературы появится позже.