
The association of inflammatory status and immunological parameters with single-nucleotide polymorphisms of cytokine and Toll-like receptor genes in patients with schizophrenia
Aннотация
Background: Schizophrenia is a multifactorial and clinically polymorphic disease with a significant hereditary component. The study of the influence of functionally significant polymorphisms of the genes regulating immune response and inflammation on the immune parameters in patients with schizophrenia is relevant for the development of new methods for predicting the course of the disease and finding new therapy targets. The aim of the study: To study the association of polymorphisms in cytokine and Toll-like receptor genes with the serum levels of immune mediators in patients with schizophrenia. Materials and methods: 95 patients diagnosed with schizophrenia and 99 healthy volunteers were recruited into the study. Polymorphisms of genes regulating the immune response (IL1B rs1143627, IL4 rs2243250, IL2 rs2069762, IL10 (rs1800896, rs1800872, rs1800871), IFNL rs8099917, IFNL rs12979860, TLR9 rs5743836, TLR9 rs352140, TLR3 rs3775291), the serum level of major cytokines and markers of systemic inflammation were determined. Results: The patients were found to have elevated levels of key pro-inflammatory cytokines IL-8 and IL-17A and other markers of systemic inflammation, as well as an increase in the content of antigen-antibody complexes in the bloodstream. Increased frequency of IL10 gene polymorphisms (rs1800896, rs1800872, rs1800871) was revealed in the patients with schizophrenia. A relationship between IL-6 and IL-8 levels and the carriage of IL1B T-511C (rs16944), IL10 1082A (rs1800896), IL10 592 C>A (rs1800872) and TLR3 Leu412Phe (G/A) (rs3775291) single nucleotide polymorphisms was found. Conclusion: The revealed associations may indicate a role of SNPs in cytokine and Toll-like receptor genes in modulating the network genetic interactions underlying certain immunological endophenotypes in the patients. Based on the results of this work and literature data, it is important to further study the relationship of immunogenetic markers in schizophrenia with the immune profiles of the disease and its clinical manifestations, including in larger samples of patients.
Ключевые слова: cytokines, inflammation, single nucleotide polymorphisms, schizophrenia, Toll-like receptors
К сожалению, текст статьи доступен только на Английском
Introduction. Neuroimmune interactions mediate homeostasis and normal functioning of both the central nervous system (CNS) and the immune system with the participation of neurotransmitters, hormones and cytokines produced in the cells of these systems. Changes in the signal pathways that provide cooperation between the immune system and the nervous system are involved in the pathogenesis of a number of non-infectious diseases, including mental disorders [1].
Schizophrenia is a multifactorial disease with a significant hereditary component (more than 60% of the disease risk according to twin studies). There is evidence showing the critical role of immune disorders and neuroinflammation in the pathogenesis of various forms of schizophrenia [2]. In recent years, a number of works confirm the role of systemic inflammation in the development of this disease [3]. Numerous clinical, postmortem, biomedical, genetic and experimental studies, including cell and animal models, have shown that schizophrenia is accompanied by neuroinflammation and systemic immune disorders [4, 5, 6]. According to the results of large genome-wide association studies (GWAS), single-nucleotide polymorphisms in genes regulating the functions of the immune system have significant associations with schizophrenia risk [7, 8]. Patients with psychotic episode have increased levels of inflammation markers in the cerebrospinal fluid and systemic circulation, and an increased expression of proteins involved in the immune response and inflammation in the prefrontal cortex [4-7]. According to indirect evidence, schizophrenia associated with neuroinflammation can account for up to 40% of cases, but more convincing studies are needed [2].
Maes et al., who studied the protein-protein interaction network in patients with first-episode schizophrenia, have shown that schizophrenia is characterized by prominent changes in the nuclear factor-κB signaling related to inflammation, in Toll-like receptor 4 (TLR4) signaling mediating the immune response to bacterial infections and in JAK/STAT (Janus kinases/signal transducer and activator of transcription) pathways mediating the effects of cytokines [8]. According to the authors, these data suggest that schizophrenia is a systemic neuro-immune disorder that is triggered by infectious stimuli and induces neuro-immune toxicity cascades overexpressed in people that have hereditary predisposition such as reduced anti-inflammatory protection and altered Toll-like receptor signaling. It is now generally accepted that the genetic component of the risk of schizophrenia is determined by the contribution of polymorphisms of many genes, with each polymorphism having only a small effect on hereditary predisposition. Moreover, all known genetic markers of schizophrenia in total provide only less than 50% of the family predisposition which is actually observed in the population according to the data of twin and genealogical studies, indicating the important role of the environment in the realization of hereditary risk [9].
Polymorphisms of the genes regulating the immune response are among the most important genetic risk factors of schizophrenia [10-17]. Some of the strongest associations with the risk of the disease are observed for the major histocompatibility complex (MHC) locus, and, in particular, for the complement component C4 gene located within this locus [11]. In addition, results of GWAS studies indicate a significant relationship with the risk of schizophrenia for at least 39 genes that regulate the immune response and are located outside the MHC locus. More than 30 of these genes are expressed in both the brain and immune cells [18]. It was revealed that a number of immunogenetic markers (at least 12 out of 39 markers outside the MHC locus) are common for schizophrenia and other mental and neurological diseases, including bipolar disorder, anxiety disorder and amyotrophic lateral sclerosis [19]. Common genetic markers in schizophrenia and immune-mediated diseases including Crohn's disease, multiple sclerosis, rheumatoid arthritis, systemic lupus erythematosus, type 1 diabetes mellitus, and ulcerative colitis have also been found [20]. Interestingly, according to a GWAS study, most of the polymorphisms that occur with the highest frequency in schizophrenia affect the signal pathways specific to the functioning of B cells [21].
Schizophrenia is characterized by heterogeneity of symptoms, responses to treatment and outcomes, reflecting the existence of different subtypes of the disease, but clinical observations alone generally do not allow the identification of etiological and pathogenetic differences between these subtypes [22]. It has been demonstrated in a number of studies that immune disturbances are associated with severity and type of symptoms in the patients with schizophrenia. For example, the serum level of an important marker of systemic inflammation, IL-6, has been shown to correlate with the severity of cognitive symptoms in outpatients with schizophrenia (r = -0.395) [23]. According to another study, the level of IL-6 decreased to normal values in hospitalized patients with schizophrenia after a course of antipsychotic therapy, and the decrease in IL-6 level correlated with a decrease in the positive and negative symptoms according to the PANSS scale [24]. In addition, there is evidence that an increase in the serum concentration of this cytokine is associated with morphometric changes in left lingual and left fusiform gyrus in patients with schizophrenia [25].
To search for new prognostic markers and targets for therapy, it is necessary to identify the causes of the development of immunological changes in schizophrenia, and the research should include investigation of the genetic basis of various types of immune disorders in the patients. However, the associations of genetic factors with immune parameters in the patients with schizophrenia have not been studied sufficiently [26, 27].
The aim of the study was to study the possible associations of cytokine and Toll-like receptor gene polymorphisms with the immune parameters in patients with schizophrenia for the subsequent development of new prognostic methods and identification of new therapy targets.
Materials and Methods. 95 patients with paranoid schizophrenia were enrolled into the study. 56 of the patients were men and 29 of the patients were women, the average age of the patients was 26.77 ± 1.26 years. At the time of hospitalization, the patients had signs of Kandinsky-Clerambo syndrome: delusions of influence, pseudohallucinations and mental automatism. The diagnosis was confirmed by the compliance with the diagnostic criteria of schizophrenia (F20) of the International Classification of Diseases of the tenth revision (ICD-10). The demographic and clinical characteristics of the patients and of the volunteers of the control group are presented in Table 1.

Blood sampling was done in the first days of hospitalization, after achieving reduction of the positive symptoms by antipsychotic therapy. The control group for the genetic study included 99 healthy volunteers, comparable to patients with schizophrenia in terms of gender and age, mean age 28.34 ± 2.56 years (53 women and 46 men). Exclusion criteria included somatic diseases in the phase of decompensation, signs of alcohol abuse and/or other substance use disorders, mental diseases other than schizophrenia, exacerbation of infectious, inflammatory and autoimmune diseases in the 2 months preceding the examination, pregnancy.
The study was approved by the local ethics committee of the National Research Center "Kurchatov Institute" (No. 5, 04/05/2017). All participants were familiarized with the details of the study and signed a voluntary informed consent sheet.
DNA extraction was performed using a set of reagents for RNA / DNA extraction from clinical material “AmpliPrime RIBO-prep” (AmpliSens, Moscow) according to the manufacturer's recommendations. This method is based on lysis of cells and denaturation of cellular proteins using a lysis solution containing a chaotropic agent (guanidine thiocyanate), followed by precipitation of nucleic acids with isopropanol and their further extraction into the solution.
Determination of single nucleotide polymorphisms in IFNL, IL1B, IL2, IL4, TLR3, and TLR9 genes was carried out using reagent kits for polymerase chain reaction (PCR) analysis produced by “DNA-technology” and “Litekh” according to the manufacturer's recommendations.
To identify polymorphisms of IL1B (rs1143627 T-31C) and IL4 (rs2243250 C589T) genes, we used the method of polymerase chain reaction with a fluorescent detection scheme in “real time”. With a sample of isolated DNA, two amplification reactions were carried out in parallel with two pairs of allele-specific primers. The mixture contained an intercalating dye SYBR Green, the fluorescence intensity of which multiplies upon incorporation into the resulting double-stranded product.
To identify polymorphisms of the interferon λ (IFNL) gene (rs8099917 T> G, rs12979860 C> T), the polymerase chain reaction method with a fluorescent detection scheme in “real time” was used. Signaling probes containing fluorescent labels Fam and Hex were introduced into the mixture for amplification for each variant of the genetic polymorphisms to be detected. After the PCR, thermal melting of the duplexes was carried out, the change of fluorescence level was recorded and presented in the form of a graph. If the signaling probe was partially complementary to the target DNA, the melting temperature of such a duplex was lower than in the case of complete complementarity of the probe. The analysis of the results was carried out on the basis of the melting temperature of the signal probes.
IL2 rs2069762 T-330G, IL10 rs1800896 G-1082A, IL10 rs1800872 С-592А, IL10 rs1800871 C-819T, TLR9 rs5743836 T-1237C, TLR9 rs352140 G2848A, and TLR3 rs3775291 Leu412Phe single nucleotide polymorphisms were determined by PCR with electrophoretic detection of amplification products. Two amplification reactions were carried out in parallel with two pairs of allele-specific primers. Separation of PCR amplification products was carried out in 3% agarose gel. To visualize the PCR products in the gel, ethidium bromide solution was used as a dye.
Immunological studies were carried out in all patients of the main group and in 35 volunteers in the control group (mean age 27.1 ± 1.6 years; 13 men; 17 women). The systemic level of inflammation markers and the content of the main cytokines were determined by enzyme-linked immunosorbent assay (ELISA) using sets of reagents to detect human interferon-γ (IFNγ), human interleukin-1β (IL-1β), human interleukin-6, human interleukin-8, human interleukin-10, human interleukin-17, human tumor necrosis factor-α (TNFα) ( “Cytokin”, Russia, St. Petersburg), a set of reagents for detection of cortisol in blood serum (“HEMA”, Russia), a set of reagents for the detection of C-reactive protein (CRP) in blood serum (“HEMA”, Russia), a set of reagents for the detection of circulating immune complexes (CIC) in blood serum ("CHEMA“, Russia).
The software Excel (Microsoft, 2010) and STATISTICA 10 (Stat Soft, 2010) were used for statistical processing. The Shapiro-Wilk test was used to assess the normality of distribution. The results of immunological tests (continuous variables) were presented as medians with an interquartile range; when 2 groups were compared, the significance of differences was assessed using the Mann-Whitney test; when multiple groups were compared, the Kruskell-Wallis test was used with post-hoc pair-wise comparisons conducted using the Mann-Whitney test. The results of the clinical assessment (continuous variables) were presented as means ± 95% confidence interval and the Student’s t-test was used to assess the significance of differences. For discrete variables, Fisher’s exact test was used to assess the significance of differences if one of the expected values in contingency table was below 10, otherwise chi-squared test was used. The differences between the indicators were considered to be statistically significant at p <0.05. Benjamini-Hochberg procedure was used to correct for false discovery rate (FDR = 0.05). To assess the correlations, the Pearson correlation coefficient was used.
Results and Discussion
The levels of cytokines of innate and adaptive immunity and of systemic inflammation markers in patients with schizophrenia and in the control group.
The serum levels of inflammation markers, including C-reactive protein and circulating immune complexes, and of cortisol was significantly higher in the patients with schizophrenia than in healthy volunteers (Table 2). The level of the proinflammatory cytokine/chemokine IL-8/CCL8 was significantly higher in the patients than in the control group (p <0,05). The assessment of the level of the pro-inflammatory cytokines IL-1β and TNFα has shown there were no significant differences between patients with schizophrenia and healthy volunteers (Table 3). Although the difference in the levels of IL-6, an important inflammatory cytokine, between the patients and the control group was statistically significant, showing an increase in the content of IL-6 in the patients, the statistical power did not reach 0.8, so larger samples are needed to confirm this finding.
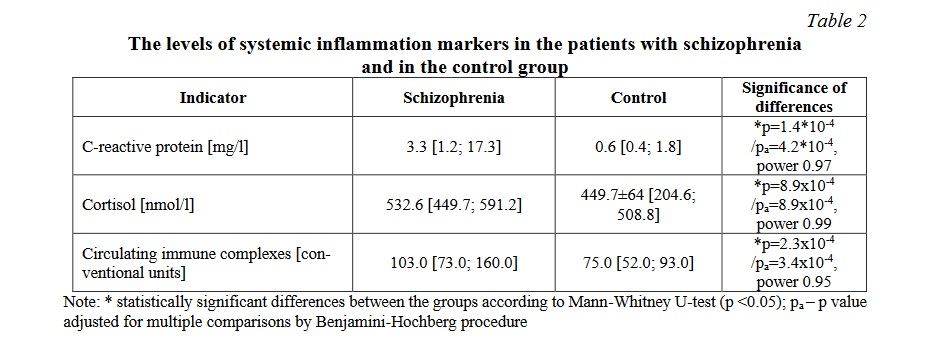

These findings indicate that in schizophrenia there was an activation of the systemic inflammatory response and of the pituitary-hypothalamus-adrenal axis. An increase in the CIC content may indicate the activation of the formation of antigen-antibody complexes in the bloodstream, as well as disruption of their clearance.
Given the presence of systemic inflammation, including increased levels of IL-8, we also assessed the level of a key immunoregulatory cytokine, IL-10, in patients with schizophrenia. Its central function is to suppress inflammation and excessive immune activation. In the patient group, the content of this cytokine was increased (more than 5 pg/ml) in 20 out of 30 patients, and in the control group it increased only in three people; however, the differences turned out to be insignificant, which could be associated with the high variability of the IL-10 level in schizophrenic patients. In one third of the patients with normal IL-10 concentration, the content of IL-6 or IL-8 or IL-1β was increased, which indicates suppression of immunoregulatory mechanisms in these patients.
The serum levels of a key Th2 cytokine, IL-4, did not differ significantly between the groups (Table 3). The level of IFNγ, an important Th1 cytokine, was increased (more than 45 pg/ml) in 53 of 87 patients with schizophrenia and only in 6 of 19 healthy volunteers, and its level was significantly higher in schizophrenia (p <0.05), but the statistical power of the comparison did not reach 0.8, so larger samples are needed to confirm this finding. An increase in the level of IL-17A above 1 pg/ml was observed significantly more often in schizophrenia: in 35 out of 73 patients with schizophrenia and only in 1 out of 16 people of the control group (p <0.05 according to Fisher's exact test). IL-17A is the most important cytokine of the Th17 response, involved in the stimulation and maintenance of inflammation and autoimmune responses. It is implicated in the pathogenesis of neurodegenerative diseases including Alzheimer's disease, Parkinson's disease, multiple sclerosis, amyotrophic lateral sclerosis, and glaucoma [28]. Of 35 patients with an increase in IL-17 levels (more than 1 pg/ml), only 7 had normal IFNγ level (below 45 pg/ml). Thus, a significant proportion of schizophrenic patients had signs of excessive activation of the Th17 and Th1 responses.
Polymorphisms of cytokine and Toll-like receptor genes in patients with schizophrenia and in the control group.
The frequency of the studied polymorphisms was checked for compliance with the Hardy-Weinberg equilibrium. It was found that the Hardy-Weinberg law is fulfilled for all the studied polymorphisms.
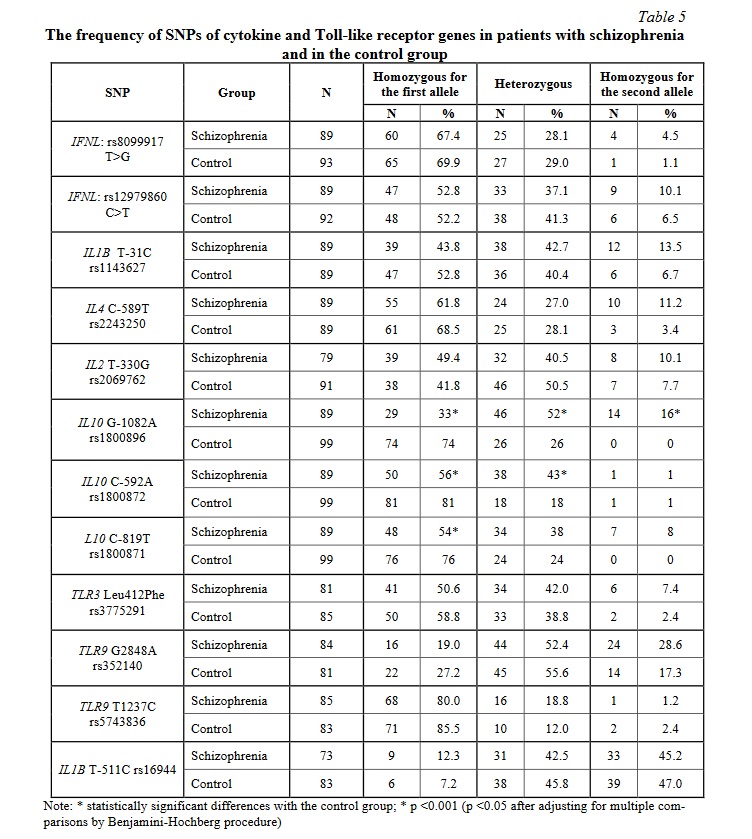
Statistically significant differences in the frequency of SNPs of the IL10 gene between the patients and the control group were found (Table 5). The frequency of homozygous state for the first allele of all studied polymorphisms of the IL10 gene was significantly (p <0.001) lower in the patients than in the control group (OR = 0.16, 95% CI [0.09; 0.31], power 0.99 for IL10 G-1082A rs1800896; OR = 0.28, 95% CI [0.15; 0.55], power 0.95 for IL10 С-592А rs1800872; OR = 0.35, 95% CI [0.19; 0.66], power 0.85 for IL10 C-819T rs1800871). Moreover, the frequency of heterozygous state for the G-1082A (rs1800896) and C-592A (rs1800872) polymorphisms was significantly (p <0.001) higher in the patients: OR = 3.95, 95% CI [2.13; 7.30], power 0.94 for IL10 G-1082A rs1800896; OR = 3.69, 95% CI [1.90; 7.16], power 0.95 for IL10 С-592А rs1800872. The frequency of homozygous state for the second allele for IL10 G-1082A rs1800896 was also significantly (p <0.001) higher in the patient group: OR = 38.22, 95% CI [2.24; 650.92], power 0.99. No significant differences between the patients with schizophrenia and the control group were found in the frequency of the other studied SNPs of cytokine and Toll-like receptor genes (Table 4). However, there was a tendency for higher frequency of the homozygous state for the minor allele of most of the studied SNPs in the patients with schizophrenia.
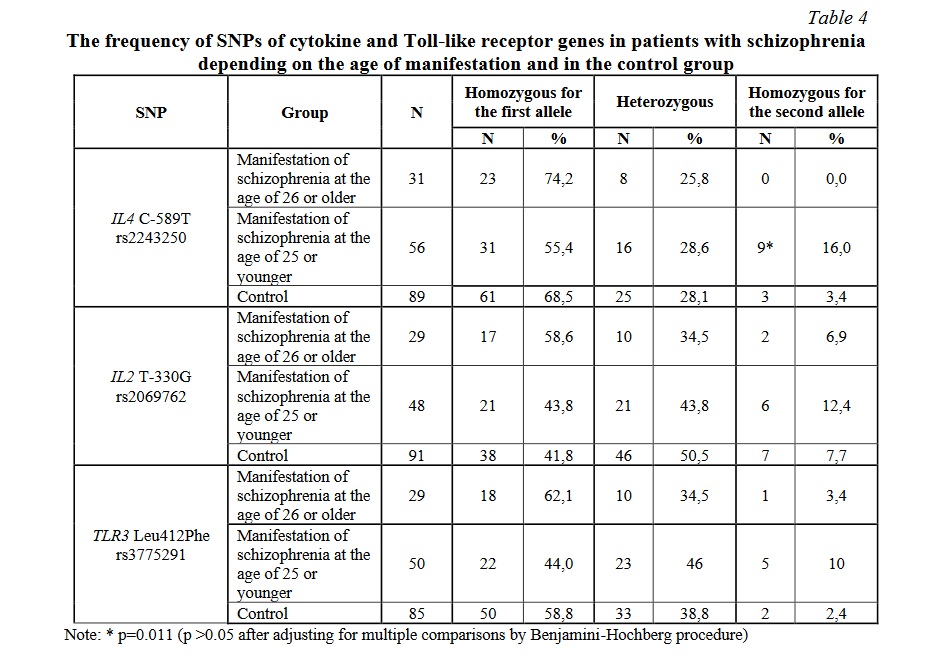
The analysis of associations between the frequency of SNPs, socio-demographic data and clinical characteristics of the patients has revealed that homozygous state for the T allele of C-589T (rs2243250) SNP of the IL4 gene was more common in the patients in whom the disease manifested at the age of 25 years or less (on average, 19.02 ± 1.02 years) compared with the control group: OR = 5.49, 95% CI [1.42; 21.36]; p = 0.011. However, the statistical power (0.67) did not reach 0.8, so larger samples are needed to confirm this finding. In the patients in whom schizophrenia manifested at the age of more than 25 years (on average, 30.38 ± 1.24 years), there were no differences in the frequency of the studied SNPs with the control group (Table 5).
Associations of single-nucleotide polymorphisms of cytokine and Toll-like receptor genes with serum levels of immune mediators in patients with schizophrenia.
The following new associations between single-nucleotide polymorphisms of cytokine and Toll-like receptor genes and serum levels of immune mediators were revealed in patients with schizophrenia.
It was found that carriage of the C allele for the IL1B T-511C polymorphism (rs16944) was associated with an increase in the level of IL-6 in patients with schizophrenia (p=0.034 for differences with the control group, power 0.94). In the patients who were homozygous for the T allele of this SNP the level of IL-6 did not differ from the control group (Figure 1).
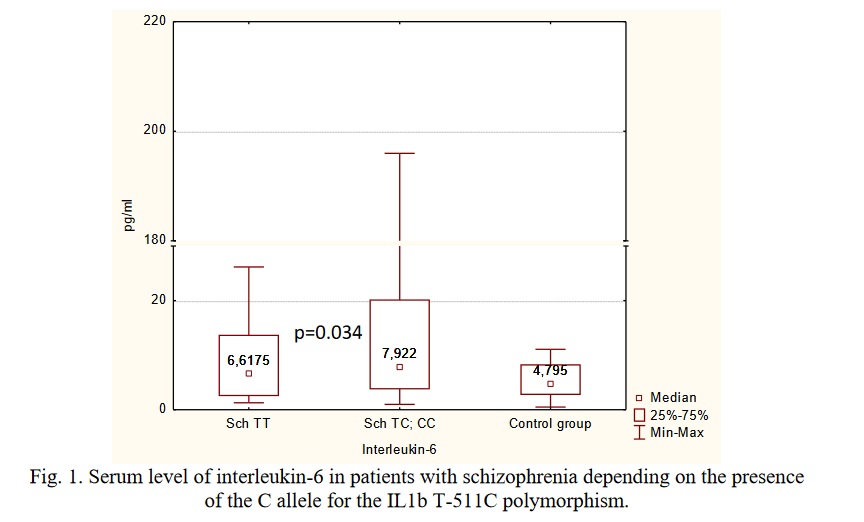
According to the literature, the IL1B T-511C polymorphism influences the transcription of the IL1B gene, but the results in the literature regarding the functional effects of different alleles are not consistent. According to [29], the T allele results in increased transcriptional activity of the ILB gene in the THP-1 human monocyte cell line. Yet, there is data that in the carriers of GG genotype the peripheral blood mononuclear cells release more IL-1β after stimulation with lipopolysaccharide [30]. Importantly, the GG genotype for the IL1B T-511C polymorphism has been associated with an increased risk of schizophrenia, odds ratio 1.24 (95% CI 1.09, 1.41) [31]. It has also been shown that the rs16944 GG genotype is associated with decreased gray matter volume and functional activity of the brain in schizophrenia and bipolar disorder [33, 33]. In addition, this genotype was associated with a reduced likelihood of achieving remission in 256 individuals with depression (odds ratio = 1.74) [34].
The mechanisms of the influence of the IL1B T-511C polymorphism on the risk of schizophrenia and on the changes of the brain in the patients remain unclear, but it has been hypothesized that the effect of this SNP is realized through modulation of IL-1β secretion levels [35]. In the pathogenesis of schizophrenia, chronic activation of innate immunity and disrupted balance in activation of Th1 and Th2 links of adaptive immunity with excessive activation of Th2 link plays a large part, contributing to an increase in the levels of kynurenic acid that causes glutamatergic dysfunction in the patients [36]. IL-1β is one of the key inflammatory cytokines, involved in the initiation and sustainment of chronic inflammation. Activating the NFκB signaling, it induces expression of a number of inflammatory mediators in target cells, including the inflammatory cytokine IL-6 [37]. IL-6 promotes Th2 differentiation and inhibits Th1 differentiation through two independent molecular mechanisms. IL-6 activates nuclear factor of activated T cells (NFAT), stimulating production of IL-4 by CD4(+) T cells and their differentiation into Th2 cells. Moreover, it upregulates suppressor of cytokine signaling (SOCS)-1 to inhibit IFNγ signaling and the differentiation of Th1 cells [38].
The role of IL-6 as a potential marker of the progression of cognitive impairment and brain MRI changes in schizophrenia is currently being intensively studied. As mentioned above, serum IL-6 levels are negatively associated with cognitive performance in schizophrenia [26]. In addition, according to our previous results, there is evidence that an increase in the levels of this cytokine is associated with morphometric changes in the left lingual and left fusiform gyrus in the patients [25]. Taking this into account, the association of the IL1B T-511C polymorphism with an increase in IL-6 levels may contribute to the pathogenetic mechanisms of schizophrenia at least in some of the patients. Further research is needed to understand the possible effect of the IL1B T-511C polymorphism on the clinical characteristics of the disease and on its prognosis.
Carriage of the homozygous G allele of the IL10 SNP 1082A (rs1800896) was associated with no significant increase in the levels of proinflammatory cytokine/chemokine IL-8 (29.08 [12.22; 101.54], while carriers of the A allele had significantly increased IL-8 levels compared with the control group (39.07 [14.33; 89.67] and 13.16 [8.33; 32.11], p = 0.027, power 0.99).
Carriage of the A allele of the IL10 SNP 592 C>A (rs1800872) was also associated with no significant increase in the levels of IL-8 (29.08 [9.24; 55.23], while homozygotes for the C allele had significantly increased IL-8 levels compared with the control group (51.62 [23.36; 106.24] and 13.16 [8.33; 32.11], p = 0.02, power 0.99).
According to the literature, homozygous carriage of the G allele for the IL-10 polymorphism G-1082A is associated with a high level of IL-10 production [39, 40]. The highest IL-10 production is detected in the carriers of the G allele which also have the A allele in IL10 592 C>A SNP [40]. IL-10 is an immunoregulatory cytokine that inhibits the production of inflammatory mediators including IL-8, which may to a certain extent explain the lack of increased IL-8 levels in the patients with schizophrenia who are carriers of IL10 1082A and IL10 592 C>A SNPs.
Carriage of the Phe allele for the TLR3 Leu412Phe (G / A) (rs3775291) polymorphism was also associated with no significant increase in the levels of IL-8 compared to the control group, while carriers of the Leu allele had significantly increased levels of IL-8, p<0,05, power 0.89 (Figure 2). As can be seen from Figure 2, carriers of the Leu allele also had significantly higher levels of TNFα than in the control group, but the statistical power of this comparison was lower than 0.8, therefore larger studies are required to confirm the result.
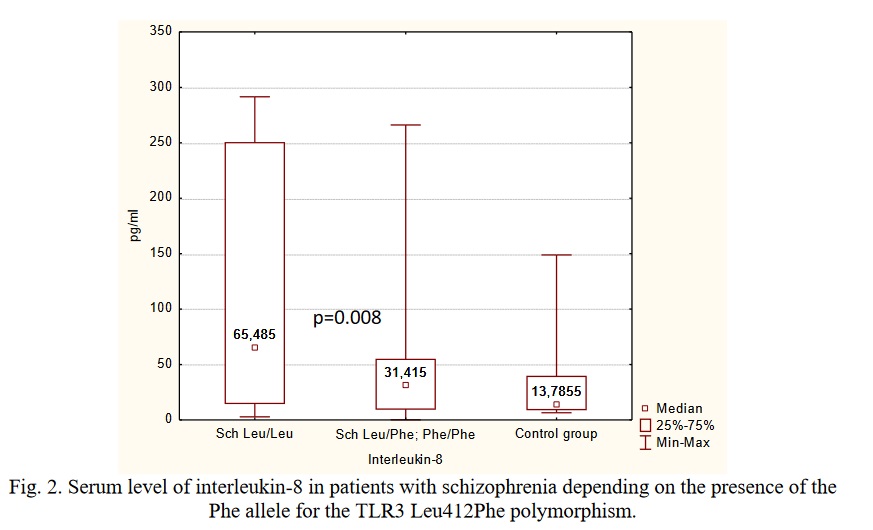
According to the literature, increased serum levels of IL-8 in schizophrenia predict the development of resistance to therapy [41]. Further studies are needed to elucidate the clinical characteristics and the course of the disease in the patients with schizophrenia who carry the SNPs associated with lower IL-8 levels.
There is evidence that the TLR3 Leu412Phe (G/A) genetic polymorphism (rs3775291) reduces the ability of the TLR3 protein to bind double-stranded RNA (dsRNA). This polymorphism is associated with a decrease in the synthesis of IFNγ by mononuclear cells of the umbilical cord blood after stimulation of the cells with a synthetic TLR3 ligand, pI:C [42]. Therefore, the polymorphism may contribute to a less pronounced activation of the innate immune response and a lower activation of the Th1-link of adaptive immunity. In addition, it has been demonstrated to have some protective effect against the risk of HIV infection and HSV-2 infection [43, 44]. In accordance with these effects of the SNP, our work shows a lower level of proinflammatory cytokines in the patients with schizophrenia who are carriers of the TLR3 Leu412Phe polymorphism. They also demonstrate an increase in the CIC level which may reflect a higher activation of the Th2 component of adaptive immunity.
In the present study a number of associations were revealed between the polymorphisms in the genes regulating the immune response and the parameters of the systemic immunity in patients with schizophrenia. The new data obtained are important for describing different immunological variants of schizophrenia and for identifying genetic factors that affect the type and severity of immunological disorders in patients.
Activation of the systemic inflammatory response found in this study in patients with schizophrenia is in accordance with the literature data and with the previous results obtained by the authors [25].
Increased frequency of SNPs in the IL10 gene in patients with schizophrenia has been revealed in this study. This finding indicates a need of further studies to clarify the possible role of IL10 SNPs in the disruptions of neuro-immune interactions that contribute to the pathogenesis of schizophrenia and might affect the clinical manifestations of the disease.
Associations of SNPs in the genes regulating the immune response with the immune parameters in schizophrenia were also found. Notably, such associations were identified for SNPs that were overrepresented in the patients, including IL10 1082A (rs1800896) and IL10 592 C>A (rs1800872). These results may indicate a role of these SNPs in modulating the network genetic interactions underlying certain immunological endophenotypes in the patients. The associations of SNPs in genes regulating the immune response with increased production of key inflammatory cytokines such as IL-6 and IL-8 warrants further research, considering that level of IL-8 has been shown in our previous work to correlate with clinically important characteristics of the disease including the severity of negative symptoms, and level of IL-6 was shown to be associated with structural changes of the brain detected by MRI [25, 45].
Based on the results of this study, researching the associations of polymorphisms of genes regulating the immune response with immune parameters and clinical characteristics of patients with schizophrenia is a promising approach to elucidating the pathogenetic mechanisms of the disease. Further research, particularly in larger samples of patients, adopting big data and network methodologies, may contribute to the translation of fundamental immunogenetic data into clinical practice as well as to finding new markers for prediction of the course of schizophrenia.
Conclusion. In this study, the parameters of immunity and a number of SNPs of cytokine and TLR genes in patients with schizophrenia have been assessed. The associations of the studied SNPs with the type and severity of immune disorders and the presence of systemic inflammation in patients with schizophrenia have been analyzed. New data have been obtained on the relationship of genetic factors, immune disorders and systemic inflammation in schizophrenia.
The principal findings of the study are:
1. In the patients with schizophrenia, activation of the systemic inflammatory response with an increase in the levels of key pro-inflammatory mediators including C-reactive protein, cortisol, circulating immune complexes, IL-8 and IL-17A was revealed, which is in accordance with the previous results obtained by the authors and with the literature.
2. Increased frequency of IL10 gene polymorphisms (rs1800896, rs1800872, rs1800871) was revealed in the patients with schizophrenia compared to the control group.
3. New associations between the levels of immunological and inflammatory markers and SNPs of cytokine and TLR genes in patients with schizophrenia were found. Increased IL-6 levels were associated in the patients with the presence of the C allele for IL1B T-511C (rs16944), and increased IL-8 levels were associated in the patients with the presence of the G allele for IL10 SNP 1082A (rs1800896), the A allele for IL10 592 C>A (rs1800872) and the Phe allele for TLR3 Leu412Phe (G/A) (rs3775291).
Financial support
This work was supported by NRC “Kurchatov Institute” (R&D "Biomedical Technologies", subtopic "Immunogenetic strategies for the diagnosis and treatment of schizophrenia", order No. 1059 of July 2, 2020, order No. 2757 of October 28, 2021) and partially supported by RSF (grant 20-15-00299).














Список литературы
Kerage D, Sloan EK, Mattarollo SR, et al. Interaction of neurotransmitters and neurochemicals with lymphocytes. Journal of Neuroimmunology. 2019;332:99-111. DOI: https://doi.org/10.1016/j.jneuroim.2019.04.006
Bechter K. Updating the mild encephalitis hypothesis of schizophrenia. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2013;42:71-91. DOI: https://doi.org/10.1016/j.pnpbp.2012.06.019
Müller N, Krause D, Weidinger E, et al. Immunological treatment options for schizophrenia. Fortschritte der Neurologie-Psychiatrie. 2014;82(4)210-219. Deutsch. DOI: https://doi.org/10.1055/s-0033-1355776
Goldsmith DR, Rapaport MH, Miller BJ. A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Molecular Psychiatry. 2016;21(12):1696-1709. DOI: https://doi.org/10.1038/mp.2016.3
Miller BJ, Buckley P, Seabolt W, et al. Meta-analysis of cytokine alterations in schizophrenia: clinical status and antipsychotic effects. Biological Psychiatry. 2011;70(7):663-671. DOI: https://doi.org/10.1016/j.biopsych.2011.04.013
Potvin S, Stip E, Sepehry AA, et al. Inflammatory cytokine alterations in schizophrenia: a systematic quantitative review. Biological Psychiatry. 2008;63(8):801-808. DOI: https://doi.org/10.1016/j.biopsych.2007.09.024
Malashenkova IK, Krynskiy SA, Ogurtsov DP, et al. Immunoinflammatory profile in patients with episodic and continuous paranoid schizophrenia. Consortium Psychiatricum. 2021;2(1):19-31. DOI: https://doi.org/10.17816/CP66
Maes M, Plaimas K, Suratanee A, et al. First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection. Cells. 2021;10(11):2929. DOI: https://doi.org/10.3390/cells10112929
Agerbo E, Sullivan PF, Vilhjálmsson BJ, et al. Polygenic Risk Score, Parental Socioeconomic Status, Family History of Psychiatric Disorders, and the Risk for Schizophrenia: A Danish Population-Based Study and Meta-analysis. JAMA Psychiatry. 2015;72(7):635-641. DOI: https://doi.org/10.1001/jamapsychiatry.2015.0346
Ruzzo EK, Geschwind DH. Schizophrenia genetics complements its mechanistic understanding. Nature Neuroscience. 2016;19(4):523-525. DOI: https://doi.org/10.1038/nn.4277
Sekar A, Bialas AR, de Rivera H, et al. Schizophrenia risk from complex variation of complement component 4. Nature. 2016;530(7589):177-183. DOI: https://doi.org/10.1038/nature16549
Schafer DP, Lehrman EK, Kautzman AG, et al. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012;74(4):691-705. DOI: https://doi.org/10.1016/j.neuron.2012.03.026
Dickerson F, Boronow J, Stallings C, et al. The lymphotoxin Cys13Arg polymorphism and cognitive functioning in individuals with schizophrenia. Schizophrenia Research. 2007;89(1-3):173-176. DOI: https://doi.org/10.1016/j.schres.2006.08.015
Li X, Zhang W, Lencz T, et al. Common variants of IRF3 conferring risk of schizophrenia. Journal of Psychiatric Research. 2015;64:67-73. DOI: https://doi.org/10.1016/j.jpsychires.2015.03.008
Srinivas L, Vellichirammal NN, Alex AM, et al. Pro-inflammatory cytokines and their epistatic interactions in genetic susceptibility to schizophrenia. Journal of Neuroinflammation. 2016;13(1):105. DOI: https://doi.org/10.1186/s12974-016-0569-8
García-Bueno B, Gassó P, MacDowell KS, et al. Evidence of Activation of the Toll-like Receptor-4 Proinflammatory Pathway in Patients with Schizophrenia. Journal of Psychiatry and Neuroscience. 2016;41(3):E46-E55. DOI: https://doi.org/10.1503/jpn.150195
Mak M, Misiak B, Frydecka D, et al. Polymorphisms in immune-inflammatory response genes and the risk of deficit schizophrenia. Schizophrenia Research. 2018;193:359-363. DOI: https://doi.org/10.1016/j.schres.2017.06.050
Pouget JG. The Emerging Immunogenetic Architecture of Schizophrenia. Schizophrenia Bulletin. 2018;44(5):993-1004. DOI: https://doi.org/10.1093/schbul/sby038
MacArthur J, Bowler E, Cerezo M, et al. The new NHGRI-EBI Catalog of published genome-wide association studies (GWAS Catalog). Nucleic Acids Research. 2017;45(D1):D896-D901. DOI: https://doi.org/10.1093/nar/gkw1133
Wang Q, Yang C, Gelernter J, et al. Pervasive pleiotropy between psychiatric disorders and immune disorders revealed by integrative analysis of multiple GWAS. Human Genetics. 2015;134:1195-1209. DOI: https://doi.org/10.1007/s00439-015-1596-8
Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511(7510):421-427. DOI: https://doi.org/10.1038/nature13595
Bhati MT. Defining Psychosis: The Evolution of Schizophrenic Spectrum Disorders DSM-5. Current Psychiatry Reports. 2013;15(110):409. DOI: https://doi.org/10.1007/s11920-013-0409-9
Ribeiro-Santos R, de Campos-Carli SM, Ferretjans R, et al. The association of cognitive performance and IL-6 levels in schizophrenia is influenced by age and antipsychotic treatment. Nordic Journal of Psychiatry. 2020;74(3):187-193. DOI: https://doi.org/10.1080/08039488.2019.1688389
Luo Y, He H, Zhang J, et al. Changes in serum TNF-α, IL-18, and IL-6 concentrations in patients with chronic schizophrenia at admission and at discharge. Comprehensive Psychiatry. 2019;90:82-87. DOI: https://doi.org/10.1016/j.comppsych.2019.01.003
Malashenkova IK, Ushakov VL, Krynskiy SA, et al. The relationship of morphometric changes of the brain with IL-6 levels, systemic inflammation and immune disturbances in the patients with schizophrenia. Procedia Computer Science. 2021;190:553-559. DOI: https://doi.org/10.1016/j.procs.2021.06.064
International Schizophrenia Consortium, Purcell SM, Wray NR, et al. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature. 2009;460(7256):748-752. DOI: https://doi.org/10.1038/nature08185
Schizophrenia Psychiatric Genome-Wide Association Study Consortium. Genome-wide association study identifies five new schizophrenia loci. Nature Genetics. 2011;43(10):969-976. DOI: https://doi.org/10.1038/ng.940
Chen J, Liu X, Zhong Y. Interleukin-17A: The Key Cytokine in Neurodegenerative Diseases. Frontiers in Aging Neuroscience. 2020;12:566922. DOI: https://doi.org/10.3389/fnagi.2020.566922
Chen H, Wilkins LM, Aziz N, et al. Single nucleotide polymorphisms in the human interleukin-1B gene affect transcription according to haplotype context. Human Molecular Genetics. 2006;15(4):519-529. DOI: https://doi.org/10.1093/hmg/ddi469
Iacoviello L, Di Castelnuovo A, Gattone M, et al. Polymorphisms of the interleukin-1beta gene affect the risk of myocardial infarction and ischemic stroke at young age and the response of mononuclear cells to stimulation in vitro. Arteriosclerosis, Thrombosis, and Vascular Biology. 2005;25:222-227. DOI: https://doi.org/10.1161/01.ATV.0000150039.60906.02
Shirts BH, Wood J, Yolken RH, et al. Association study of IL10, IL1β, and IL1RN and schizophrenia using tag SNPs from a comprehensive database: Suggestive association with rs16944 at IL1β. Schizophrenia Research. 2006;88(1-3):235-244. DOI: https://doi.org/10.1016/j.schres.2006.06.037
Fatjó-Vilas M, Pomarol-Clotet E, Salvador R, et al. Effect of the interleukin-1β gene on dorsolateral prefrontal cortex function in schizophrenia: a genetic neuroimaging study. Biological Psychiatry. 2012;72(9):758-765. DOI: https://doi.org/10.1016/j.biopsych.2012.04.035
Papiol S, Molina V, Desco M, et al. Gray matter deficits in bipolar disorder are associated with genetic variability at interleukin-1 beta gene (2q13). Genes, Brain and Behavior. 2008;7(7):796-801. DOI: https://doi.org/10.1111/j.1601-183X.2008.00421.x
Baune BT, Dannlowski U, Domschke K, et al. The interleukin 1 beta (IL1B) gene is associated with failure to achieve remission and impaired emotion processing in major depression. Biological Psychiatry. 2010;67(6):543-549. DOI: https://doi.org/10.1016/j.biopsych.2009.11.004
Sasayama D, Hori H, Teraishi T, et al. Possible association between interleukin-1β gene and schizophrenia in a Japanese population. Behavioral and Brain Functions. 2011;7:35. DOI: https://doi.org/10.1186/1744-9081-7-35
Chiang SS, Riedel M, Schwarz M, et al. Is T-helper type 2 shift schizophrenia-specific? Primary results from a comparison of related psychiatric disorders and healthy controls. Psychiatry and Clinical Neurosciences. 2013;67(4):228-236. DOI: https://doi.org/10.1111/pcn.12040
Weber A, Wasiliew P, Kracht M. Interleukin-1 (IL-1) pathway. Science Signaling. 2010;3(105):cm1. DOI: https://doi.org/10.1126/scisignal.3105cm1
Diehl S, Rincón M. The two faces of IL-6 on Th1/Th2 differentiation. Molecular Immunology. 2002;39(9):531-6. DOI: https://doi.org/10.1016/s0161-5890(02)00210-9
Cardona-Castro N, Sánchez-Jiménez M, Rojas W, et al. IL-10 gene promoter polymorphisms and leprosy in a Colombian population sample. Biomedica. 2012;32(1):71-6. DOI: https://doi.org/10.1590/S0120-41572012000100009
Schotte H, Schlüter B, Schmidt H, et al. Putative IL-10 Low Producer Genotypes Are Associated with a Favourable Etanercept Response in Patients with Rheumatoid Arthritis. PLoS ONE. 2015;10(6):e0130907. DOI: https://doi.org/10.1371/journal.pone.0130907
Enache D, Nikkheslat N, Fathalla D, et al. Peripheral immune markers and antipsychotic non-response in psychosis. Schizophrenia Research. 2021;230:1-8. DOI: https://doi.org/10.1016/j.schres.2020.12.020
Cho P, Gelinas L, Corbett NP, et al. Association of common single-nucleotide polymorphisms in innate immune genes with differences in TLR-induced cytokine production in neonates. Genes and Immunity. 2013;14(4):199-211. DOI: https://doi.org/10.1038/gene.2013.5
Huik K, Avi R, Pauskar M, et al. Association between TLR3 rs3775291 and resistance to HIV among highly exposed Caucasian intravenous drug users. Infection, Genetics and Evolution. 2013;20:78-82. DOI: https://doi.org/10.1016/j.meegid.2013.08.008
Svensson A, Tunbäck P, Nordström I, et al. Polymorphisms in Toll-like receptor 3 confer natural resistance to human herpes simplex virus type 2 infection. Journal of General Virology. 2012;93(Pt 8):1717-1724. DOI: https://doi.org/10.1099/vir.0.042572-0
Malashenkova IK, Ushakov VL, Zakharova NV, et al. Neuro-immune aspects of schizophrenia with severe negative symptoms: new diagnostic markers of disease phenotype. Sovremennye Tehnologii v Medicine. 2021;13(6):24-35. DOI: https://doi.org/10.17691/stm2021.13.6.03