
Studies to elucidate the effect and antiapoptotic mechanism of 2-ethyl-3-hydroxy-6-methylpyridine-n-acetyltaurinate in a rat model of retinal ischemia–reperfusion
Aннотация
Background: The study of possible ways of effective correction of retinal ischemia-reperfusion injury, which accompanies a number of eye diseases, is relevant today. The aim of the study: To study the retinoprotective effect and antiapoptotic mechanism of 2-Ethyl-3-hydroxy-6-methyl-pyridine-N-acetyltaurinate (EHMP-NAT) in a rat model of retinal ischemia–reperfusion (I/R). Materials and methods: A pathology model with an increase in intraocular pressure (IOP) to 110 mmHg was used. The retinoprotective effect of EHMP-NAT at a dose of 4.4 mg/kg/day, in comparison with emoxipine and taurine in equimolar doses, was estimated by the changes in the retinal microcirculation, electroretinograms (the b/a coefficient), and retinal caspase-3, NF-κB p65, p53 gene expressions in Wistar rats. Results: The use of EHMP-NAT led to an increase in the retinal microcirculation level to 756.5 (median) perfusion units in comparison with emoxipine (p = 0.045) and taurine (p = 0.00029). The b/a coefficient increased in comparison with the group with emoxipine (p = 0.0099) and with the group with taurine (p = 0.015). In the group with EHMP-NAT, the caspase-3 gene expression decreased reliably in comparison with emoxipine (p = 0.0002) and with taurine (p = 0.0028); the NF-κB p65 gene expression decreased in comparison with emoxipine (p = 0.0009) and with taurine (p = 0.0022); the p 53 gene expression decreased in comparison with emoxipine (p = 0.0022) and with taurine (p = 0.0009). Conclusion: Based on the data obtained, in correction of retinal I/R by EHMP-NAT, improvements in the retinal microcirculation, functional state, and caspase-3, NF-κB p65, p53 gene expressions were more pronounced than in monotherapy with emoxipine or taurine.
Ключевые слова: retinal ischemia–reperfusion, rats, 2-ethyl-3-hydroxy-6-methyl-pyridine-n-acetyltaurinate, apoptosis
К сожалению, текст статьи доступен только на Английском
Introduction. The retina is susceptible to lipid peroxidation because it consists of lipids with high levels of polyunsaturated fatty acids, such as docosahexaenoic acid, cis-arachidonic acid and choline phosphoglyceride. Prostanoids are synthesized from arachidonic acid as a result of the sequential action of phospholipase A2 and cyclooxygenase, which are triggered by oxidative stress and peroxidation. The accumulation of peroxides ultimately contributes to the production of thromboxane A2, which is a powerful cytotoxic agent for microvessels [1]. Platelet activation factor and lysophosphatidic acid are other lipids formed during lipid peroxidation, which act as pro-inflammatory mediators and contribute to damage to retinal microvessels. Platelet activation factor is produced in large quantities during oxidative stress, and its cytotoxic effects are largely mediated by thromboxane A2. Similarly, lysophosphatidic acid is released from lysophosphatidylcholine under the action of lysophospholipase D and may play a role in retinal inflammation, which leads to microvascular cytotoxicity [2]. Retinal microvascular injuries lead to a decrease in perfusion parameters and microcirculation.
The high rate of blood flow makes the retina susceptible to ischemia-reperfusion (I/R) injury. It is widely accepted that I/R injury is caused by the increased generation of reactive oxygen species (ROS) during the process of I/R, including superoxide (O2), hydrogen peroxide (H2O2) and hydroxyl radicals (OH-). The overproduction of ROS can induce apoptosis in the retina [3, 4, 5]. As a result, inflammation and cell apoptosis are considered to be major causes of the pathological changes following I/R injury. Induction of an inflammatory response by reperfusion may increase the intensity of damage.
Many studies have shown that after I/R injury, treatment with antiapoptotic agents is effective for preserving cell populations throughout the retina [6-9]. p53 is a transcription factor that induces apoptosis in response to DNA damage, oncogene activation, hypoxia [10]. p53 may mediate apoptosis through extrinsic and intrinsic pathways. The intrinsic pathway is associated with Bcl-2 family proteins, which controls the mitochondria outer membrane permeabilization [11, 12]. Upregulation of p53 inhibits Bcl-xL and Bcl-2 protein, contributes to the release of cytochrome c, and promotes caspase-3–mediated apoptosis [13]. NF-κB is a transcription factor activated by oxygen free radicals, an increase in its immunoactivity in retinal inner nuclear layer (INL) and ganglion cells layer after I/R injury was consistent with TUNEL-labeling apoptosis [14]. In NMDA–mediated retinal excitotoxicity, NF-κB promotes an apoptotic response in ganglion cells via p53 upregulation [15].
The research object is a new molecule 2-ethyl-3-hydroxy-6-methyl-pyridine-N-acetyltaurinate (EHMP-NAT) that includes the following pharmacophores: 3-hydroxypyridine and N-acetyltaurinate. EHMP-NAT was synthesized at the All-Russian Scientific Center for Safety of Biologically Active Substances, Russia, and is related to the class of 3-hydroxypyridines. 3-hydroxypyridine substantiates antioxidant effect and the correction of glutamate excitotoxicity [16]. As is known from the literature data, 2-ethyl-3-hydroxy-6-methylpyridine hydrochloride (Emoxipine) has an antioxidant effect due to its ability to interact with Fe2+ and radicals and to enhance efficiency of the endogenous antioxidant protection [17, 18]. In the retina, taurine is present in high concentration, regulates Ca2+ transport and has antioxidant properties [19]. Taurine also suppresses glutamate-induced Ca2+ flow [20].
The problem of search for ways to improve the effectiveness of pharmacological correction of retinal ischemia is relevant and beyond doubt [21,22]. In this experimental study, we studied the possibility of pharmacological correction with 2-ethyl-3-hydroxy-6-methyl-pyridine-N-acetyltaurinate in retinal I/R, assessing its effect on microcirculation in the retina, the functional state of the retina and the expression of pro-apoptotic factor genes in the retina.
Objective: to study the retinoprotective effect and antiapoptotic mechanism of EHMP-NAT in a rat model of retinal I/R.
Materials and Methods
1. Animals
The experiments were approved by the Local Ethics Committee of Belgorod State National Research University, Belgorod (Protocol #11/20 from 7th of September 2020). The animals were obtained from the Stolbovaya laboratory animal nursery in the Moscow region, Russia. The study was performed in 50 Wistar rats weighing 225–275 g. Ethical principles of handling laboratory rats were observed in accordance with the European Convention for the Protection of Vertebrate Animals Used for Experimental and Other Scientific Purposes, CETS No. 123. All manipulations on rats were performed under general anesthesia with i.p. administration of chloral hydrate solution.
2. Design of the Experiment
The following groups were included in the experiment:
(1) control group (an equivalent volume of water for injections was administered using the same manner as the studied substance) (n = 10);
(2) group with the retinal I/R model (n = 10);
(3) group with I/R correction by EHMP-NAT (n = 10);
(4) group with I/R correction by emoxipine (n = 10);
(5) group with I/R correction by taurine (n = 10).
A model of retinal I/R was performed under general anesthesia using i.p. chloral hydrate, 300 mg/kg, by applying mechanical pressure to the anterior eye chamber for 30 min [23]. Mechanical pressure was 110 mm Hg and carried out by an atraumatic metal rod in the cylinder with calibrated piston system. The system for retinal I/R simulation was calibrated as followed: IOP was registered by administrating into the anterior eye chamber 30G needle sensor (Biopac System, Inc., Goleta, CA, USA) with use the TSD104A hard-ware-software complex MP150 production Biopac System, Inc., and the computer pro-gram AcqKnowledge 4.2. Rendering mechanical pressure was carried out as per step calibration system of the cylinder with the registration of the increase of IOP that allowed us to estimate the level of IOP increase at a specific position.
EHMP-NAT (All‐Russian Scientific Center for Safety of Biologically Active Substances, Staraya Kupavna, Russia) was administered parabulbarly (into the orbital adipose body, the area of the lower eyelid) as a 1% solution at a dose of 4.4 mg/kg/day daily for 4 days, including the first injection 30 min before I/R.
Emoxipine (1% solution for injections, Federal State Unitary Enterprise “Moscow Endocrine Plant”, Moscow, Russia) as a comparison drug was administered at a dose of 2 mg/kg [5] in the same manner as EHMP-NAT.
Taurine (4% solution for injections, “Dalkhimpharm” JSC, Khabarovsk, Russia) as a comparison drug was administered at a dose of 1.8 mg/kg in the same manner as EHMP-NAT. The studied substance and comparison drugs were taken for research in equimolar doses.
The retinoprotective effects of pharmacological agents were estimated by changes in retinal microcirculation level using laser Doppler flowmetry (LDF), and electroretinography (ERG) after 72 hours of reperfusion [23]. The expression of proapoptotic factor genes (Caspase 3, NF-κB p65, p53) in the retina was evaluated by quantitative real-time PCR.
3. LDF
72 hours after ischemia, the microcirculation level in rats was measured by LDF. The online registration was performed using MP150 production Biopac System, Inc., a computer‐based data acquisition system with AcqKnowledge 4.2 software, and a TSD‐144 needle‐type sensor (Biopac System, Inc., Goleta, CA, USA). After the rats were anesthetized, the microcirculation level was measured at 10 points on the circumference of the eye. The duration of the recording at each point was about 20 sec. Then the average value was calculated, which was taken as the value of the retinal microcirculation level in a particular rat [23]. An example of recording the microcirculation level in the eye of a rat from the control group is presented in Figure 1.
4. ERG
ERG was performed according to the protocol previously published [8]. Evoked biopotentials were run at a frequency of 1–1000 Hz, amplified, averaged, and presented graphically on the screen using the MP150 data acquisition and analysis system (Biopac Systems, Inc., Goleta, CA, USA). ERG registration was carried out in response to a single stimulation. To assess the degree of retinal ischemia, the b/a coefficient was calculated.
5. Quantitative real-time PCR
10 to 20 mg of retinal tissue was cleaned with ice cold PBS 0.1 M solution before immersion in RNAlater Stabilisation Solution to avoid degradation of cellular RNA. Before RNA extraction, retinal tissue was sonicated in 300 µl of lysis buffer. The total RNA was extracted as follows: the samples were homogenized in a commercial solution "Extract RNA" (Evrogen, Russia) at the rate of 50 mg of tissue per 1 ml of extractor. The resulting suspension was centrifuged for 5 minutes at 15000g at room temperature. Then, the samples were washed sequentially and centrifuged with chloroform, 100% isopropanol, and 75% ethanol cooled. 15 µl of distilled water was added to the resulting sediment and vortexed. The concentration of RNA in the resulting solution was determined using a NanoDrop 2000C spectrophotometer (Thermo Scientific Inc., USA) three times. After calculating the average concentration, the samples were adjusted to an RNA content of 200 ng/ml.
For the translation of RNA into cDNA, the reverse transcription reaction was carried out in a Bio Rad CFX96 amplifier using a commercial MMLV RT kit (Evrogen, Russia) at the rate of 11 µl of the sample per 9 µl of the reverse transcription kit. Reaction temperature conditions: 60 minutes at 39 °C + 10 minutes at 70 °C. Then, to evaluate the expression of targeted genes in the Bio Rad CFX96 amplifier, PCR of samples was performed using a commercial SYBR® Green Master Mix kit (Bio-Rad Laboratories, Inc., USA) and oligonucleotide primers (Evrogen, Russia).
Primers were selected using the Primer-BLAST resource (NCBI, USA) (Table 1).

After amplification, the relative expression value for each gene was calculated using the formula:
EGOI =2^(CtACTB - ΔCtGOI),
where CtACTB is the cycle threshold value for the reference gene ACTB.
6. Statistical Analysis
In all cases, the median (Me) and lower (QL) and upper (QU) quartiles were calculated. Between‐group differences were analyzed by the Mann–Whitney U test. Statistical analyses were performed using Statistica 10.0 software.
Results
1. LDF results
In the control group, the microcirculation level was 748.5 (median; QL = 733.0; QU = 768.0) perfusion units. In the group with I/R, the microcirculation level was 357.0 (median; QL = 314.0; QU = 385.0) perfusion units, which was significantly lower (p = 0.00016) than in the control group. The use of EHMP-NAT led to an increase in this parameter to 756.5 (median; QL = 680.0; QU = 774.0) perfusion units compared to the group with retinal I/R (p = 0.00016); compared to the group with emoxipine (p = 0.045); compared to the group with taurine (p = 0.00029). In the group treated with emoxipine, the retinal microcirculation level was 685.5 (median; QL = 647.0; QU = 710.0) perfusion units, which was significantly different (p = 0.0028) from the control group, and significantly different from the group with retinal I/R (p = 0.00016). In the group with taurine, this parameter was 390.5 (median; QL = 311.0; QU = 415.0) perfusion units, which was significantly different from the control group (p = 0.00016), and didn’t differ from the group with I/R (p = 0.212).
Based on the data obtained, in the retinal I/R model, improvement in the retinal microcirculation in correction by EHMP-NAT was more pronounced than in monotherapy with emoxipine or taurine.
2. ERG results
The influences of the studied substance and comparison drugs on the a‐ and b‐wave amplitudes and the b/a coefficient in the model of retinal I/R are presented in Table 2. The a-wave amplitude values remained unchanged in all experimental groups.
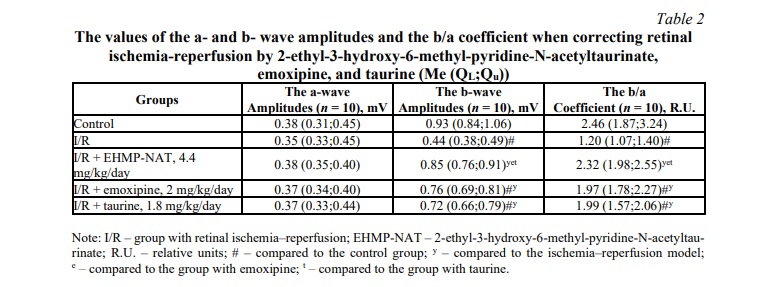
When simulating I/R, the b‐wave amplitude reduced by 2.1 times in comparison with the control group (p = 0.0002). In the group treated with EHMP-NAT, the b‐wave amplitude increased by 93.2% (p = 0.0002) in comparison with the group with I/R; the b/a coefficient increased by 93.3% in comparison with the group with I/R (p = 0.0002); by 64.2% in comparison with the group with emoxipine (p = 0.0099); by 65.8% in comparison with the group with taurine (p = 0.015). In the group with emoxipine administration, the b‐wave amplitude increased by 72.7% (p = 0.0002) in comparison with the group with I/R; the b/a coefficient increased by 64.2% (p = 0.0002) in comparison with the group with I/R. In the group with taurine, the b‐wave amplitude increased by 63.6% (p = 0.0002) in comparison with the group with I/R; the b/a coefficient increased by 65.8% (p = 0.0002) in comparison with the group with I/R.
Based on the table data from the ERG study, the best correction of functional retinal changes caused by I/R was found in group with EHMP-NAT administration.
3. Real-time PCR results
Effects of the studied substance and comparison drugs on I/R-induced changes in the retinal caspase-3, NF-κB p65, p53 genes expression were evaluated after 72 hours of reperfusion. When simulating pathology, the caspase-3 gene expression increased reliably in comparison with the control group (p = 0.00016); the NF-κB p65 gene expression in-creased by 2.5 times in comparison with the control group (p = 0.00016); the p 53 gene expression increased by 2.4 times in comparison with the control (p = 0.0002).
In the group treated with EHMP-NAT, the caspase-3 gene expression decreased by 2.3 times (p = 0.0002) in comparison with I/R group; the NF-κB p65 gene expression de-creased by 1.6 times (p = 0.0002) in comparison with the group with I/R; the p 53 gene expression decreased by 1.6 times in comparison with I/R group (p = 0.0002).
In the group with emoxipine administration, the caspase-3 gene expression decreased by 24.6% (p = 0.001) in comparison with the group with I/R; the NF-κB p65 gene expression decreased by 1.3 times (p = 0.0032) in comparison with the group with I/R; the p 53 gene expression decreased by 1.2 times in comparison with I/R group (p = 0.0008).
In the group with taurine, the caspase-3 gene expression decreased by 1.7 times (p = 0.0007) in comparison with I/R group; the NF-κB p65 gene expression was not significantly different from the group with I/R (p = 0.076); the p 53 gene expression was significantly different from the control group (p = 0.0002) and from the I/R group (p = 0.011) (Table 3).
In the group with EHMP-NAT, the caspase-3 gene expression decreased reliably in comparison with emoxipine (p = 0.0002) and with taurine (p = 0.0028); the NF-κB p65 gene expression decreased in comparison with emoxipine (p = 0.0009) and with taurine (p = 0.0022); the p 53 gene expression decreased in comparison with emoxipine (p = 0.0022) and with taurine (p = 0.0009).
Based on the table data from the real-time PCR study, the best pharmacological correction of retinal I/R by protecting retina from apoptosis was found in the group with EHMP-NAT.

Discussion. In this study, we investigated the retinoprotective effect and the mechanism of antiapoptotic action of EHMP-NAT against the retinal I/R through the evaluation of retinal microcirculation, electrophysiological state, and caspase-3, NF-κB p65, p53 gene expressions.
Several studies have shown that, in the early stage of ischemia, an increase in extracellular glutamate activates NMDA-receptors, causing a Ca2+ influx. Then, NO is produced under the NO-synthase action in the mitochondria, which leads to production of ONOO− [4,24]. Based on the literature data, glutamate receptor stimulation activates NF-κB and plays a role in ROS-induced cell apoptosis [25, 26]. NF-κB p65 has been shown to be activated in NMDA-mediated retinal excitotoxicity as well as other neurodegenerative diseases [27]. Our study demonstrated that the exposure to transient increase in IOP to 110 mmHg (ischemic episode) followed by 72-hour reperfusion increases the retinal expression of NF-κB p65 gene. It is likely that the observed changes in NF-κB expression at these time points involve response to NMDA activation in retinal glial and ganglion cells [28]. Correction with EHMP-NAT reduced NF-κB p65 gene expression in retinal I/R model.
The present study also investigated the effect of EHMP-NAT on the retinal I/R-mediated changes in the p53 and caspase-3 gene expressions. Based on the literature data, NMDA-induced retinal ganglion cell loss corresponded with increased levels of the p53 gene [8]. The inactivation or low expression of p53 provide a retinoprotective effect in retinal ischemia or excitotoxicity [29]. There is evidence that Bax and Bcl-2 expressions are highly regulated by the p53 gene [30]. Several studies have shown that, p53-mediated retinal ganglion cell apoptosis is accompanied by the activation of caspase-3 dependent signaling pathway [31, 32]. Correction with EHMP-NAT also reduced p53 and caspase-3 gene expressions in retinal I/R model.
Over the past two decades, LDF has been adapted to measure blood flow in the optic disc, subfoveal vasculature [33], and iris in humans [34]. LDF values depend linearly on the state of the blood flow. This makes it useful in assessing changes in blood flow caused by physiological or pathological stimuli that do not change the optical properties of tissues [35]. Hyperoxia, hypercapnia, flickering light stimuli, changes in ocular perfusion pressure affect blood flow in the optic disc and choroidal blood flow measured using LDF [36]. The results of our study indicate that correction with EHMP-NAT improved the retinal microcirculation against I/R.
In this study, ERG was used to confirm whether microcirculatory changes in the retina were related to functional changes. ERG has confirmatory rather than primary diagnostic value because injuries to any of the retinal layers could result in ERG changes: photoreceptors (a-wave), and the inner nuclear layer (b-wave) [37]. The amacrine and bipolar cells are located in the inner nuclear layer and are sensitive to I/R- and NMDA-mediated retinal injury [38]. As it was shown earlier, in retinal I/R, the cells of the inner nuclear layer are primarily damaged [5, 23]. The results of the present study indicate that treatment with EHMP-NAT preserved the retinal electrophysiology against I/R.
The detected retinoprotective effect of EHMP-NAT to correct experimental retinal I/R may be also associated with the following: (a) 3‐hydroxypyridine is a “scavenger” of peroxides and superoxides of fatty acids [39]; (b) taurine, an antioxidant and the predominant free amino acid in the retina that maintains membrane structural integrity, regulates Ca2+ binding and transport [40], and inhibits retinal excitotoxic injury [41].
In view of the above, further studies of the effects of EHMP-NAT on the models of NMDA‐mediated retinal excitotoxicity with intravitreal administration of NMDA, cerebral stroke, and traumatic brain injury are planned.
Conclusion. EHMP-NAT at a dose of 4.4 mg/kg of rat mass has a pronounced retinoprotective effect, which is superior to the effects of comparison drugs, emoxipine and taurine, in equimolar doses in the simulated retinal I/R. This was confirmed by the improvement in the attainment of target values of the retinal microcirculation, improvement in the functional state of the retina, downregulation of caspase-3, NF-κB p65, p53 genes expression in retina.














Список литературы
Список использованной литературы появится позже.