
Curcumin Ameliorates High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease by Regulating Endoplasmic Reticulum Stress in The Liver
Aннотация
Background: Nonalcoholic fatty liver disease (NAFLD) is a chronic pathological condition of the liver due to excess triglyceride accumulation in hepatocytes. The antioxidant properties of curcumin improve lipid dysregulation and reduce reactive oxygen species (ROS). The aim of the study: This study explores the effects of curcumin administration on beclin-1 and microtubule-associated protein light chain-3 (LC3) expression as autophagy markers and XBP1 spliced as a marker indicating endoplasmic reticulum stress in ameliorating HFD-induced NAFLD in mice. Materials and methods: Twenty-four ddY male mice were divided into four groups: the normal chow group, high-fat diet (HFD) group, HFD with curcumin 50 mg/kg for 28 days group, and HFD with curcumin 100 mg/kg for 28 days group. Bodyweight and food intake were measured daily, and curcumin was administered intraperitoneally. The animals were sacrificed 24 hours after the last treatment. The liver was collected for macroscopic and histopathological assessment and molecular analysis using the reverse transcription-polymerase chain reaction (PCR) method. Results: Curcumin 50 and 100 mg/kg improved macroscopic and histopathological features of the liver. The results of the molecular analysis showed that curcumin 50 and 100 mg/kg did not increase the beclin-1 or LC3 mRNA expression in the liver (p>0.05). Meanwhile, curcumin 100 mg/kg significantly increases the XBP1 spliced expression in the liver (p<0.05), which indicates that curcumin modulates endoplasmic stress induced-NAFLD in a dose-dependent manner. Conclusion: Curcumin overcomes endoplasmic reticulum stress in the high-fat diet-induced NAFLD in mice.
Ключевые слова: nonalcoholic fatty liver disease, curcumin, endoplasmic reticulum stress, high-fat diet, beclin-1, light chain-3, XBP1
К сожалению, текст статьи доступен только на Английском
Introduction. Nonalcoholic fatty liver disease is a spectrum of progressive liver disease which is characterized by simple steatosis, which means there are more than 5% triacylglycerol infiltration in hepatocytes, inflammation of hepatocytes, and hepatocellular ballooning due to perturbation of homeostatic mechanisms that regulate the synthesis and use of liver fat [1, 2]. NAFLD occurs in the absence of alcohol consumption or with minimal alcohol consumption [2]. NAFLD is divided into two isoforms, simple NAFL, a benign and reversible form of NAFLD, and NASH, an aggressive and irreversible form [3].
NAFLD occurs in many countries and has become a global burden, with a worldwide incidence percentage of approximately 25.24% with wide geographical variations, with the highest prevalence of NAFLD in Middle Eastern countries and South America at around 30% [4]. Meanwhile, for countries in Asia, especially in Sri Lanka, Malaysia, Singapore, and Indonesia, the prevalence of NAFLD tends to be varied, which is around 5-30% caused by wide variations of genetic differences, economic conditions, intake, and individual lifestyle [5].
The pathogenesis of NAFLD remains unclear, but there is a well-known theory, namely the two-hits theory from James and Day in 1998 [6, 7]. They defined the first hit of lipid accumulation in hepatocytes due to changes in intrahepatic lipid metabolism, then followed by a second hit in the form of a mechanism that represents the various factors that trigger the progression of NAFLD, such as inflammation of hepatocytes, inflammation, and fibrosis [6, 7]. The complexity of this process led to the assumption that the theory was too old-fashioned, so in 2010, Tilg and Moschen came up with their “multi parallel hits hypothesis” theory, which explains the pathogenesis of NAFLD more complexly; nevertheless, it seems less resonant when compared to the two hits theory by James and Day [7, 8].
Autophagy is a catabolic process that targets cell constituents such as damaged organelles into lysosomes for degradation. It regulates cell metabolism and integrates it into eliminating microorganisms to enhance inflammation and activation of the adaptive immune system [9, 10]. Autophagosome formation involves three main steps: 1) initiation of the uncoordinated 51-like kinase (ULK); 2) beclin-1/PI3K nucleation; and 3) membrane elongation with the help of LC3 [11]. Autophagy is activated in various ways, such as restriction of the rapamycin complex called mTOR and activation of protein kinase or AMPK [1]. Several proteins involved in human autophagic pathways, microtubule-associated protein light chain-3 (LC3) and beclin-1, play important roles that have been reported to be related to the physiology and pathogenesis of human liver disease [12]. Inhibition of autophagy with negative regulators interacting with beclin-1 results in accelerated lipid accumulation and triggers the pathogenesis of NAFLD [1]. LC3 is a protein that plays a role in modulating the formation of autophagosomes, in which LC3-I is formed and then conjugated to LC3-II as a marker of successful development of complete autophagosomes [13]. The downregulation of LC3 transcription is often associated with the severity of liver disease [13].
Endoplasmic reticulum stress is a mechanism that plays an essential role in the pathogenesis of NAFLD, dysregulation of the endoplasmic reticulum triggers a physiological response from cells, namely the Unfolded Protein Response, which is a compensatory mechanism for cells to restore protein folding, reduce the accumulation of misfolded proteins and maintain the integrity of the cell [14, 15]. There are three transmembrane proteins involved in this mechanism, which are inositol-requiring enzyme 1α (IRE1α), Protein Kinase R-Like ER Kinase (PERK), and activating transcription factor-6α (ATF6α) [14, 15]. X-Box Binding Protein 1, as a protein downstream of IRE1, is an imperative regulator of lipogenesis and apoptosis in cells that experience endoplasmic reticulum stress [16]. Measurement of XBP1 spliced mRNA expression is a strategic way to analyze the effect of gene expression changes in stressed condition cells related to the progression of NAFLD.
Curcumin is a polyphenolic compound found in Curcuma longa and Curcuma xanthorrhiza, Roxb, which provides various pharmacological activities, especially antioxidant [17, 18, 19]. Curcumin induces antioxidant enzymes, modulating the activity of GSH enzymes, catalase, and superoxide dismutase in neutralizing free radicals [17]. Curcumin significantly reduces oxidative stress in the liver, inhibits apoptosis, enhances cell protection, and repairs mitochondrial damage through the induction of autophagy [20]. Curcumin induces autophagy-related apoptosis in mesothelioma and chronic myelogenous leukemia K562 cells by modulating the PI3K/AKT/mTOR and NF-kB signaling pathway [21].
The aim of the study. This study explores the effects of curcumin administration on the expression of autophagy markers (beclin-1 and LC3), endoplasmic reticulum stress through XBP1 spliced mRNA expression and to discover the mechanism and correlation of these various pathways concerning the progress of NAFLD.
Materials and methods
Experimental animals
Eight-week-old ddY male mice with a healthy physical condition were used in this study. Mice were fed in individual cages measuring 20 × 20 × 15 cm and covered with 6 mm gauze with a temperature of 22 ± 2˚C and with a 12 hours light/dark cycle. Their cages were cleaned and replaced with new husks every day. All experiments were performed at the Animal Research Laboratory of the Faculty of Pharmacy, Universitas Airlangga, Surabaya, Indonesia. The protocol of this study was approved by the Ethical Committee (Animal Care and Use Committee) of the Faculty of Veterinary Medicine, Universitas Airlangga (No.2.KE.054.05.2021).
Diet, drug, and experimental design
The high-fat diet (HFD) used in this study consists of 60% beef tallow (902 kcal/100 g), which has been melted at 65˚C using a hotplate and mixed up to 100% with a normal chow diet (306.20 kcal/100 g) that has been mashed. Curcumin (Tokyo Chemical Industry Inch., Tokyo, Japan) was dissolved to 10% polysorbate 80 (vehicle) in a recenter paratus before treatment.
In this experimental study, twenty-four mice were used. All of the test animals were reared in the same way. After one week of habituation, the mice were randomly divided into four groups (n = 6) and given the same amount of food with free access to food and water. The groups included: (1) the control group which was fed with a normal chow diet and treated with 10% of polysorbate 80 for 28 days; (2) the HFD group was fed with an HFD for 28 days; (3) the low dose curcumin-treated group which was fed with HFD and curcumin 50 mg/kg i.p for 28 days; and (4) the high dose curcumin-treated group which was fed with HFD and curcumin 100mg/kg i.p for 28 days. The high-fat diet used in this work is consistent with the previous study that successfully developed the high-fat diet-induced NAFLD in mice [22]. The justification of choice doses and the administration route of curcumin are based on previous study that found curcumin 50 and 100 mg/kg exhibit antioxidant activity in the liver diseases [23, 24, 25]. At the end of the study, all mice in each group were euthanized after 28 days of treatment and 24 hours of the last treatment. The liver was extracted and put in the freezer at –80˚C for further analysis.
Liver macroscopic evaluation and histopathological examination
The macroscopic evaluation of the liver was carried out visually. Then, for histopathological examination, the liver tissues were fixed and stored in a sample container at 4 ˚C in 4% paraformaldehyde, routinely processed and embedded in paraffin, prepared into 5 μm thick sections and stained for 8 min hematoxylin-eosin (H&E), washed for 60 min in flowing tap water, counter staining at room temperature with eosin for 60 seconds. The steatosis is characterized by lipid droplets, ballooning, and inflammation on the histopathology of the liver tissue under a light microscope (magnification, ×400; Nikon Company, Tokyo, Japan).
RNA extraction and reverse transcription-polymerase chain reaction (PCR)
Measurement of beclin-1, LC3, and XBP-1 spliced (XBP1s) mRNA relative expression was obtained from the liver tissue measurement using the reverse-transcription polymerase chain reaction (PCR) method. The animals were sacrificed, and the largest lobe of the liver was taken. The purification of total RNA in the liver sample was performed with the RNA Purification Kit® (Jena Bioscience, Germany). The reverse transcription was performed using the GoScriptTM Reverse Transcription System (Promega, USA). The following primers were used: beclin-1 (forward: 5’-GAACCG CAAGATAGTGGC-3’; and reverse: 5’-CAGAGCATGGAGCAGCAA-3’), LC3 (forward: 5’-CAGGATCCATGCCGTCCCAGAAGACC-3’; and reverse: 5’-GTCCCTTTTTGCCTTGGTAG-3’), XBP1s (forward: 5’-TCTGCTGAGTCTGCAGCAG-3’; and reverse: 5’-TCTGGGGAAGGACATTTGAA-3’), and β-actin (forward: 5’-TGTTACCAACTGGGACGACA-3’; reverse: 3’-AAGGAGGCTGGAAAA GAGC-5’).
The thermal cycler process of PCR was performed using the initial denaturation for 5 minutes at 94 ˚C, 35 cycles of denaturing for 40 seconds at 94 ˚C, annealing for 1 minute at 55 ˚C, extension for 2 minutes at 72 ˚C, and final extension for 4 minutes at 72 ˚C. The separation process of aliquots from PCR was performed on 2% agarose LE (Promega, USA) using electrophoresis). Ethidium bromide (Sigma-Aldrich) was used to stain gel and photographed using UV transillumination. The ImageJ software was used to examine the band intensity.
Data analysis
The statistical analysis for the body weight and food intake data was performed using the two-way analysis of variance (ANOVA) method, followed by the Tukey’s post hoc test. Then, the analysis for the mRNA relative expressions of beclin-1, LC3, and XBP-1 spliced was performed using the one-way ANOVA method followed by the Tukey’s post hoc test (Tukey test for multiple comparisons). The data analysis was performed using GraphPad Prism 9.0.2 for Windows (Graphpad Software, San Diego).
Results
Bodyweight profile
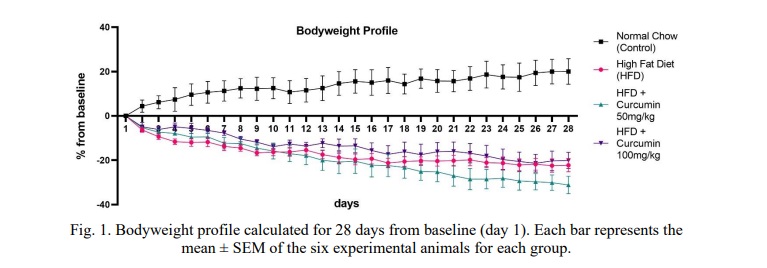
The bodyweight profiles of the normal chow group (control), the HFD-only group, and the HFD group with curcumin treatment at a dose of 50 mg/kg i.p and 100 mg/kg i.p for 28 days were compared, as shown in Fig. 1. The normal chow group tended to increase in body weight every day by 19.3% until the 28th day, inversely with the HFD group, which tended to decrease by 22.2% until the 28th day. The weight fall also occurred in the curcumin 50 and 100 mg/kg groups, which were 31.2% and 18.4% weight loss, respectively.
Food intake and calorie intake profiles
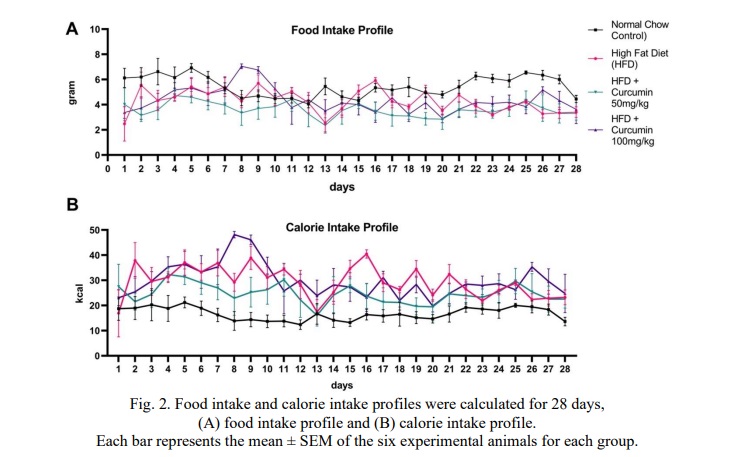
The findings showed no specific difference in the amount of food consumed by each group every day. However, in terms of calorie intake, there is a difference in the amount of calorie intake between the normal chow (control) group and the group fed with high-fat diets. The amount of calorie intake in the three groups that were fed a high-fat diet was higher compared to the normal chow group as shown in Fig. 2.
Macroscopic features of the liver
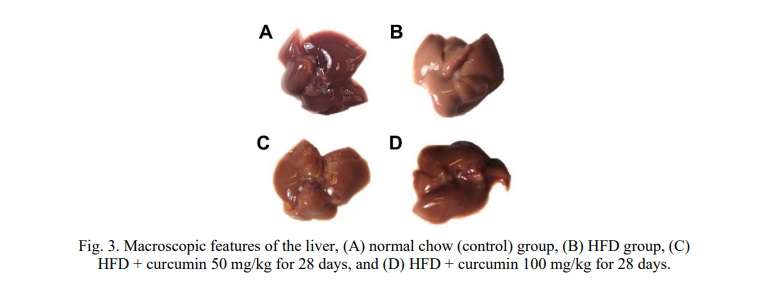
The macroscopic features of the normal liver are generally observed with many blood vessels, so the color tends to be dark red with a smooth surface, as shown in Fig. 3A (normal chow group). While in the HFD group, the liver looks pale with a granular surface due to lipid deposits, as shown in Fig. 3B. The administration of curcumin 50 or 100 mg/kg i.p for 28 days improves the macroscopic features of the liver (Fig. 3C and 3D), characterized by changes in liver color that are getting closer to normal as it becomes dark red and the surface becomes smoother compared to the HFD-only group.
Histopathological features of the liver
The histopathological features of the liver in the normal chow (control) group shows normal hepatocyte cell structure with clearly visible sinusoids and no signs of NAFLD (Fig. 4A). While in the HFD group, the histopathological features of the liver show abnormal hepatocyte cell structures with narrowed sinusoids. In addition, there are signs of NAFLD, such as lipid droplets, ballooning cells, and inflammation at a certain point (Fig. 4B). The administration of curcumin 50 and 100 mg/kg i.p for 28 days improves the histopathological features of the liver (Fig. 4C and 4D), characterized by decreases in the number of lipid droplets, ballooning cells, and inflammatory points.

LC3, beclin-1, and XBP1 spliced mRNA relative expression
Measurement of the microtubule-associated protein light chain 3 (LC3) and beclin-1 mRNA relative expression showed no significant difference between all experimental groups (Fig. 5A and 5B). Furthermore, for XBP1 spliced (XBP1s) evaluation, the findings showed a significant decrease in the XBP1s mRNA relative expression in the HFD group compared to the normal chow (control) group. The supplementation of curcumin 100 mg/kg but not 50 mg/kg increases the XBP1s mRNA relative expression compared to the HFD group (Fig. 5C).

Discussion. The high-fat diet (HFD) intake in the animal promoted liver damage, similar to the phenotype in humans with NAFLD [26]. Therefore, HFD induction is often used for in vivo NAFLD model development [26, 27, 28]. In this study, an analysis of the effect of curcumin in the repair of NAFLD was carried out from cellular properties assessment by histopathological and molecular evaluation through the evaluation of the biomarkers of autophagy and endoplasmic reticulum stress.
The administration of the HFD triggered gradual weight loss in the HFD group, HFD + curcumin 50 mg/kg group, and HFD + curcumin 100 mg/kg group. This result is in accordance with the previous study that found a weight loss in mice received HFD beef tallow 60%, HFD beef tallow 45%, and vegetable ghee due to compensation for polyunsaturated fatty acid (PUFA) deficiency regarding high saturated fatty acid (SFA) content in HFD so that the rate of lipolysis in adipose tissue is greater than its esterification [27].
The result of the HFD-induced NAFLD model in this study is similar to the ketogenic diet in terms of its effect on body weight. This finding may be due to the HFD and ketogenic diets containing HFD composition, even though the HFD-induced NAFLD model in this study did not apply carbohydrate restriction. High-fat and low-carbohydrate intakes significantly reduce insulin secretion and cause changes in metabolism through two processes, namely gluconeogenesis and ketogenesis, resulting in the conversion of protein and fat as energy sources [29]. Hence, the ketogenic diet improves glycemic control but possesses a risk of hyperlipidemia, hyperinsulinemia, elevated liver enzymes, and the development of diseases associated with NASH [30, 31].
Macroscopically, the color of normal liver is generally dark red, and the surface texture is smooth. Meanwhile, the color of the liver with NAFLD is pale with a granular surface due to the accumulation of excess lipid droplets and had a larger size, as is characteristic of the HFD group in this study. This study's findings align with previous studies, which revealed changes in the visual appearance of the liver: it became paler with increasing time of exposure to HFD [32]. The administration of curcumin on HFD-induced NAFLD showed macroscopic liver improvement, characterized by changes in color and texture closer to normal liver than the HFD group without curcumin. Previous research that explores the supplementation of hot water extract from turmeric rhizome to HFD mice with 8 weeks of treatment reduces the serum concentration of triglycerides and total cholesterol [33].
The histopathological features of the liver in NAFLD condition are characterized by lipid droplets (steatosis), hepatocellular ballooning, and lobular and portal inflammation until fibrosis [34]. In the normal chow (control) group, normal hepatocyte structure was founded with clear sinusoids. In contrast, in the HFD group, there were many lipid droplets, ballooning, and several points of inflammation. These results indicated that NAFLD successfully occurred in the HFD group. In the HFD group treated with curcumin 50 mg/kg, there were some lipid droplets and ballooning in the cells. Meanwhile, in the HFD group treated with curcumin 100 mg/kg i.p., only ballooning was found in cells. This finding shows that the administration of curcumin repairs the liver tissue damage caused by fat accumulation in the liver.
Furthermore, the reverse transcription-PCR measurements showed no significant differences regarding the LC3 and beclin-1 mRNA in the HFD with low-dose and high-dose curcumin groups. This study's results illustrated that curcumin had no effect on autophagy markers. This finding may be due to a shorter durationof the HFD, so the NAFLD model that was successfully developed is still relatively mild. There is also the possibility that each marker of the autophagosome phase works independently since many pathways induce and inhibit autophagy mechanisms. NAFLD/NASH caused by HFD is highly variable, most likely influenced by diet composition and duration as well as animal species, strain, sex, and age [31].
Regarding the mechanism of endoplasmic reticulum stress, the HFD increases the activity of the three main transmembrane proteins IRE1, PERK, and ATF6, and causes a physiological response called unfolded protein response, which is under normal conditions, the activity of transmembrane proteins is at a bare minimum [14]. Many factors trigger endoplasmic reticulum stress, including nutritional deficiencies, accumulation of free fatty acids, hyperlipidemia, and hyperglycemia, including the induction with HFD that triggers an increase in misfolded protein in the endoplasmic reticulum lumen [14]. Along with an increase in misfolded protein, there is a decrease in the affinity of GRP78 or BiP with transmembrane proteins so that GRP78 binds to misfolded proteins in the ER lumen [14, 35]. There was an increase in misfolded protein in the lumen of the endoplasmic reticulum in cells under stress conditions, so GRP78 or BiP, which have functions to bind proteins in the endoplasmic reticulum lumen, was unable to balance the amount of misfolded protein and causes transautophosphorylation of IRE1 [14, 35]. In the IRE1 branch, transautophosphorylation of IRE1 occurred as a corollary of HSP47 release from the protein domain. The proximal sensor of endoribonuclease IRE1α induces unconventional splicing of XBP1 mRNA to develop a mature mRNA encoding an active transcription factor XBP1s afterward [14, 35].
XBP1 is a downstream gene from IRE1 in the RNA kinase branch, which has two isoforms, namely XBP1 unspliced (XBP1u) and XBP1 spliced (XBP1s) [36]. Under physiological conditions or conditions without endoplasmic reticulum stress, XBP1u expression predominates [36, 37]. Activation due to endoplasmic reticulum stress caused splicing of XBP1u by excluding 26 nucleotides resulting in a frameshift from the XBP1 coding sequence and then translated into XBP1s protein, which then translocates into the nucleus and providing changes in cells that affect lipogenesis, apoptosis, and so on [36, 38]
The reverse transcription-PCR evaluation showed a significant decrease of XBP1s mRNA relative expression in the HFD group compared to the normal chow group. Although there was an increase in splicing XBP1, this phenomenon happened in the early phase. After XBP1s increase even to supraphysiological levels, a gradual decrease appeared in the expression of spliced XBP1, which induced the activation of the PERK-ATF4 pathway and caused a terminal UPR response that resulted in an impact on cell apoptosis [39, 40]. This phenomenon underlies the decrease in the ratio of XBP1 spliced in stress condition cells in this study. The finding in this study is in accordance with the previous study that found the upregulation of IRE1 in mouse embryonic fibroblasts 2 (MEF2) cells up to 4 folds and also XBP1 splicing in WT cells up to 80% by induction of thapsigargin and tunicamycin as endoplasmic reticulum inducers, then decreased gradually [41].
In the curcumin group, there was a significant increase of XBP1s mRNA relative expression that was proportional to the increase of curcumin dose to the HFD group; it was because curcumin showed activity in modulating endoplasmic reticulum stress. Curcumin plays a role in the downregulation of GRP78, eIF2, and CHOP, a proapoptotic cell. Hence, curcumin suppresses cell apoptosis [21].
Conclusion. Curcumin improves the macroscopic and histopathological features of the liver of the HFD-induced NAFLD model by increasing the XBP1s mRNA relative expression. However, further research is still needed to explore the overall mechanisms regarding the ameliorating effects of curcumin on NAFLD.
Financial support
This research was funded by the Ministry of Education, Culture, Research and Technology of the Republic of Indonesia.














Список литературы
Список использованной литературы появится позже.