
Оценка вклада межлокусных взаимодействий полиморфных локусов генов матриксных металлопротеиназ и генов фолатного цикла материнского организма в формирование веса новорожденного
Aннотация
Актуальность: Вес ребенка при рождении является важным индикатором нормального протекания беременности. Высокий/низкий вес новорожденного является фактором риска нарушений здоровья (включая смертность) во взрослом возрасте. Цель исследования: Изучить вклад межлокусных взаимодействий полиморфных локусов генов матриксных металлопротеиназ и генов фолатного цикла материнского организма в формирование веса новорожденного. Материалы и методы: На выборке беременных (n=691) проведено молекулярно-генетическое исследование 5 полиморфных локусов генов матриксных металлопротеиназ (rs1799750 [1G>2G] MMP1, rs243865 [C>T] MMP2, rs3025058 [6A>5A] MMP3, rs11568818 [T>C] MMP7, rs17577 [G>A] MMP9) и 5 полиморфных локусов генов фолатного цикла (rs1805087 [A>G] MTR, rs1801394 [G>A] MTRR, rs1979277 [C>T] SHMT1, rs699517 [C>T] TYMS, rs2790 [A>G] TYMS). Межлокусные взаимодействия изученных SNP, ассоциированных с весом новорожденного, были изучены с использованием модификации метода снижения размерности (MDR) – MB-MDR (Model Based Multifactor Dimensionality Reduction). Для вес-ассоциированных SNP in silico были оценены их функциональные эффекты. Результаты: По результатам исследования было выявлено 9 наилучших моделей межгенных взаимодействий, связанных с весом новорожденного, в которые вошли все 10 изученных SNP (pperm≤0,001). Максимальное количество моделей было установлено для rs11568818 MMP7 (5 моделей) и rs1979277 SHMT1 (6 моделей). 4-х локусная модель rs1979277 SHMT1×rs11568818 MMP7×rs3025058 MMP3×rs1799750 MMP1 демонстрирует наиболее значимую ассоциацию с весом новорожденного. Заключение: Таким образом, межлокусные взаимодействия полиморфных локусов генов матриксных металлопротеиназ и генов фолатного цикла материнского организма ассоциированы с весом новорожденного
Ключевые слова: полиморфизм, ассоциации, межгенные взаимодействия, гены матриксных металлопротеиназ, гены фолатного цикла, вес новорожденного
Введение. Росто-весовые параметры плода находятся в прямой зависимости от оптимального процесса плацентации, поскольку плацента выполняет жизненно важные функции, включая транспорт веществ, питание, выделение, эндокринную регуляцию [1]. Новорожденные с высоким или низким весом имеют повышенный риск возникновения нарушений здоровья (включая смертность) в течение жизни в сравнении с детьми со средними значениями массы тела [1]. Физиологическая беременность у человека характеризуется постоянным ремоделированием коллагенового внеклеточного матрикса, что позволяет адаптировать плодные оболочки и матку к росту плода [2]. Неполная инвазия трофобласта и нарушение ремоделирования спиральных артерий матки вызывают постоянную ишемию и гипоксию плаценты, что увеличивает риск развития задержки роста плода (ЗРП) и других неблагоприятных исходов беременности [3-6].
Одной из важных групп регуляторов ангиогенеза и ремоделирования матки являются матриксные металлопротеиназы (ММП) [7]. ММП представляют собой семейство гомологичных цинкзависимых эндопептидаз, которые классифицируются на основе их структуры и функции как коллагеназы, желатиназы, стромелизины, матрилизины и ММП мембранного типа [8, 9]. По мере развития беременности ММП участвуют в имплантации бластоцисты, ремоделировании спиральных артерий и формировании плаценты [9]. Вследствие этого, аномальная активность металлопротеиназ нарушает нормальный процесс плацентации, способствуя развитию ЗРП [10-16].
Помимо этого, важную роль в развитии плода и плаценты играет так называемый одноуглеродный метаболизм, одним из звеньев которого является обмен фолиевой кислоты и метионина [17]. Фолатный и метиониновый циклы способствуют синтезу доноров метильных групп, позволяя клеткам использовать их для биосинтеза ДНК и пуринов, а также в реакциях метилирования ДНК, РНК, гистонов [18]. Метилирование, в свою очередь, обеспечивает множество клеточных процессов, имеющих ключевое значение в процессах имплантации, апоптоза во время органогенеза и общего внутриутробного развития плода [18]. Повышенная/пониженная активность ферментов фолатного цикла, может приводить к дефициту фолиевой кислоты и гипергомоцистеинемии [19]. При высоких концентрациях гомоцистеин вызывает воспаление, приводит к окислительному стрессу и связан с повышенным риском осложнений беременности [19-22].
Стоит отметить, что на данный момент существует достаточно малое количество исследований, в которых изучали связь между генетическими полиморфизмами матриксных металлопротеиназ и весом новорожденного/ЗРП [23]. Основная часть данных работ направлена на изучение генетических факторов преэклампсии в различных мировых популяциях [24-27]. Также ряд работ демонстрируют, что полиморфизмы в генах, кодирующих ферменты фолатного цикла, могут влиять на превращение гомоцистеина в метионин, что в итоге приводит к повышению концентрации гомоцистеина в крови и клетках, повреждению ДНК и возможным неблагоприятным исходам беременности, таким как преэклампсия и задержка роста плода [28-31].
Таким образом, обобщая данные литературных источников, можно сказать, что на данный момент существует ограниченное количество исследований, посвященных изучению роли генов-кандидатов в формирование росто-весовых показателей новорожденного. При этом результаты таких работ часто характеризуются неоднозначностью и слабой воспроизводимостью в различных популяциях мира. Это обуславливает потребность в проведении дальнейших исследований генетических факторов, влияющих на вес и рост новорожденного в популяциях Российской Федерации.
Цель исследования. Изучить вклад межлокусных взаимодействий полиморфных локусов генов матриксных металлопротеиназ и генов фолатного цикла материнского организма в формирование веса новорожденного.
Материалы и методы исследования. В наше исследование были включены женщины (n=691), которые наблюдались в течение беременности на базе перинатального центра областной клинической больницы г. Белгорода с 2008 по 2016 гг. При включении женщин в выборку применялись следующие критерии включения: 1) подписали добровольное информированное согласие для участия в исследовании; 2) являются неродственными индивидами русской национальности, родившиеся в Центрально-Чернозёмном регионе РФ; 3) наличие одноплодной беременности на сроке 36-41 недель, завершившейся живорождением. Не участвовали в исследовании женщины согласно следующим критериям исключения: 1) отказ от подписания добровольного информированного согласия; 2) родственные связи различной степени между участниками; 3) нерусский этнос; 4) место рождения, отличное от Центрально-Чернозёмного региона РФ; 5) многоплодная беременность; 6) аномалии развития матки, пуповины или плода; 7) роды, произошедшие до 36 недели беременности.
Методом фенол-хлороформной экстракции было произведено выделение геномной ДНК из венозной крови. Для последующего генотипирования был проведен отбор 10 однонуклеотидных полиморфизма (SNP) генов матриксных металлопротеиназ (rs1799750 [1G>2G] MMP1, rs243865 [C>T] MMP2, rs3025058 [6A>5A] MMP3, rs11568818 [T>C] MMP7, rs17577 [G>A] MMP9) и генов фолатного цикла (rs1805087 [A>G] MTR, rs1801394 [G>A] MTRR, rs1979277 [C>T] SHMT1, rs699517 [C>T] TYMS, rs2790 [A>G] TYMS) с учетом следующих критериев: 1) полиморфизмы генов, белковые продукты которых имеют важное значение для внутриутробного развития эмбриона и плода (согласно информации из баз данных KEGG PATHWAY [32], Reactome Pathway [33]); 2) наличие функциональных эффектов: несинонимические замены, эпигенетическая роль, связь с экспрессией и альтернативным сплайсингом генов (онлайн-ресурс Haploreg 4.2 [34]); 3) частота минорного аллеля в европейской популяции составляет ≥ 5%. Для генетического анализа отобранных SNP использовалась разновидность метода полимеразной цепной реакции (ПЦР) – ПЦР в режиме реального времени (real-time ПЦР). Для выявления генотипов применялись специфические флуоресцентные красители (TaqMan зонды). Реакция ПЦР проводилась на амплификаторе CFX96 (Bio-Rad, США).
Статистический анализ. Для всех исследованных SNP генов-кандидатов была проведена сравнительная оценка между наблюдаемым распределением аллелей/генотипов и ожидаемым по закону Харди-Вайнберга. При оценке распределения для учёта количества исследованных полиморфизмов (n = 10) применялась поправка Бонферрони (pbonf ≥ 0,005 (0,05/10). При расчетах использовали ковариаты (возраст женщины, индекс массы тела (ИМТ) до беременности, наличие осложнений беременности (задержка роста плода, преэклампсия и их сочетание), уровень артериального давления, возраст наступления менархе, количество беременностей и количество абортов). Помимо этого, для минимизации риска получения ложноположительных результатов при расчетах учитывалась поправка на множественные сравнения (осуществлялся адаптивный пермутационный тест с расчётом рpermutation (pperm)).
Так как распределение веса новорождённых в изученной выборке не было нормальным (оценено с помощью критерия Шапиро-Уилка) для проведения дальнейшего анализа в программной среде R были рассчитаны трансформированные значения данных параметров [35]. Для моделирования ассоциированных с весом ребенка при рождении межлокусных взаимодействий была использована модификация метода снижения размерности (MDR) – MB-MDR (Model Based Multifactor Dimensionality Reduction) (https://github.com/imbs-hl/mbmdR, дата обращения 18 сентября 2025 года). Были оценены двух-, трех- и четырехлокусные модели. Для визуализации межгенных сетей взаимодействия в виде энтропийного графа применялся метод снижения размерности MDR (Multifactor Dimensionality Reduction) (http://www.multifactordimensionalityreduction.org/, дата обращения 18 сентября 2025 года) [36].
Для вес-ассоциированных полиморфных локусов и SNP, находящихся с ними в сильном сцеплении (коэффициент корреляции r2≥0,80), оценивали их функциональное значение. Для решения этой задачи мы использовали шесть современных биоинформатических инструментов [37]: HaploReg [34] – эпигенетические изменения; SIFT (Sorting Tolerant From Intolerant) [38] и PolyPhen 2 (Polymorphism Phenotyping v2) [39] – несинонимичные мутации; Blood eQTL browser [40] и GTExportal [41] – оценка влияния SNP на экспрессию и уровень альтернативного сплайсинга генов в различных органах и тканях.
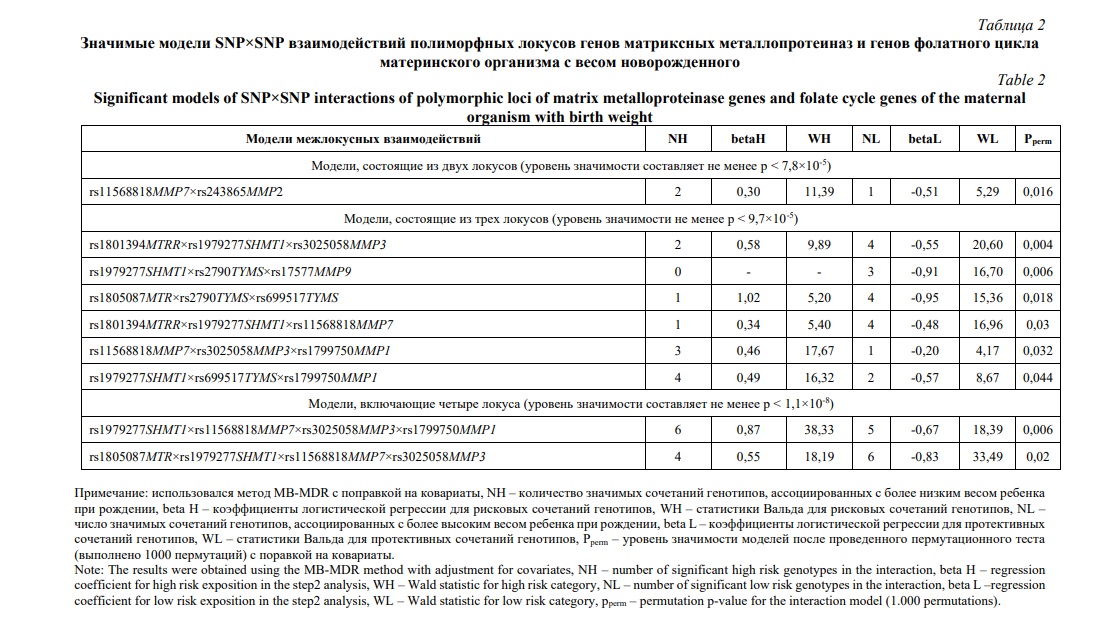
Результаты и их обсуждение. Исследование распределения частот генотипов и аллелей полиморфных локусов генов ММП и генов фолатного цикла, показало, что оно соответствуют ожидаемым значениям в соответствии с законом Харди-Вайнберга (HWE). Результаты анализа представлены в таблице 1. Также, для всех изученных SNP частота минорного аллеля (MAF) была > 5%. В рамках исследования было выявлено 9 наилучших моделей межгенных взаимодействий, связанных с весом новорожденного, в которые вошли все 10 изученных SNP (Табл. 2). Из них 1 модель была 2-х локусной, 6 моделей включали три локуса и 2 модели были 4-х локусными (pperm ≤0,001). Два полиморфных локуса rs11568818 MMP7 и rs1979277 SHMT1 входят в 5 и 6 моделей, соответственно, что соответствует наибольшему количеству моделей из всех SNP. Помимо этого, SNP-SNP взаимодействия rs11568818 MMP7 и rs1979277 SHMT1 вошли в 3 из 9 лучших моделей. Среди 9 выявленных моделей наилучшей оказалась 4-х локусная модель rs1979277 SHMT1×rs11568818 MMP7×rs3025058 MMP3×rs1799750 MMP1 (максимальный показатель Вальда (WH = 38,33, pperm = 0,006) (Рис. 1). Также, для комбинации генотипов 3-х локусной модели rs1979277CT×rs2790GA×rs17577AA была установлена наиболее значимая ассоциация с весом новорожденного (beta = -2,12, p = 0,003).
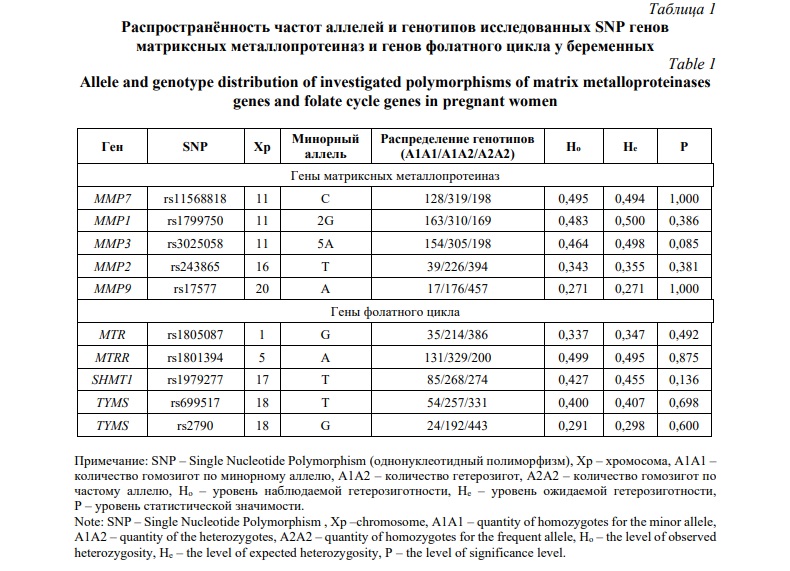
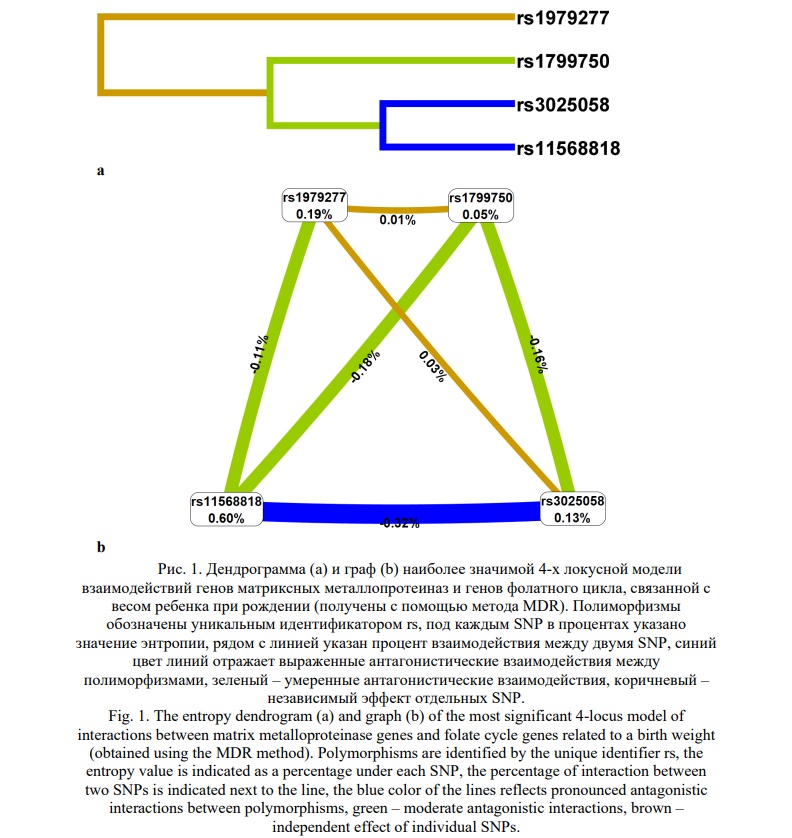
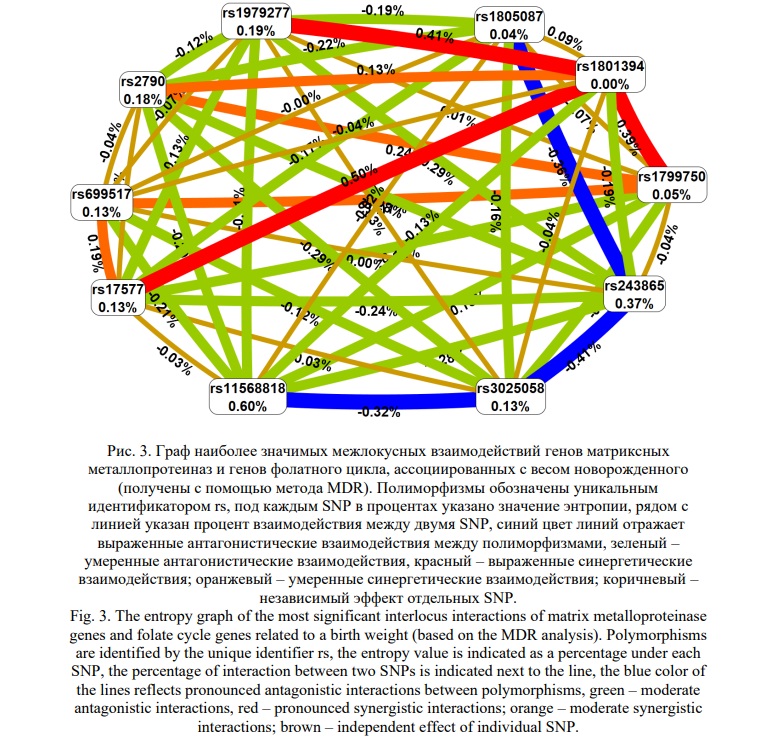
На следующем этапе анализа был применён метод MDR для оценки взаимодействия 10 однонуклеотидных полиморфизмов, связанных с весом новорожденного. Согласно дендрограмме (Рис. 2) и графу (Рис. 3), взаимодействие между изученными локусами имеет преимущественно синергетический и антагонистический характер. Синергетическое взаимодействие rs1801394 MTRR с rs1799750 MMP1, rs1979277 SHMT1 и rs17577 MMP9 было наиболее выраженным (процент энтропии 0,39%, 0,41%, 0,50%, соответственно). Кроме того, антагонистическое взаимодействие между rs11568818 MMP7, rs3025058 MMP3, rs243865 MMP2 и rs1805087 MTR было наиболее высоким (процент энтропии от -0,32% до -0,41%). Наконец, rs11568818 MMP7 и rs243865 MMP2 показали наибольший независимый эффект (процент энтропии – 0,60%, и 0,37%, соответственно).

Оценка insilico функционального значения 10-ти вес-ассоциированных SNP и 213 сильно сцепленных с ними SNP показала, что данные полиморфные локусы за счет своих функциональных эффектов определяют вовлеченность в формирование веса новорожденного 62 генов: миссенс-мутации в 6 генах (MMP3, MMP9, MTR, MTRR, SHMT1, SMCR8), эпигенетические изменения в 16 генах (ENOSF1, MMP1, MMP2, MMP3, MMP7, MMP9, MTR, MTRR, RP11-465L10.10, RP1-178F10.1, SHMT1, SMCR7, SMCR8, TOP3A, TYMS, ZNF335), регуляция экспрессии 53 генов (ALKBH5, CCDC144B, CTD-2303H24.2, DNTTIP1, DRG2, ENOSF1, EPN2-AS1, EVPLL, FAM106A, FLII, FOXO3B, KRT16P1, KRT17P2, LGALS9C, LINC02076, LLGL1, MIEF2, MMP1, MMP10, MMP7, MMP9, MTR, MTRR, MYO15A, NCOA5, NOS2P2, PLTP, PRPSAP2, RP11-212I21.2, RP11-258F1.1, RP11-258F1.2, RP11-465L10.10, RP11-806L2.2, RP11-806L2.6, RP1-253P7.4, RP1-37N7.1, RPL13P2, SHMT1, SLC12A5, SMCR8, SNX21, SYS1, TOM1L2, TOP3A, TVP23B, TYMS, TYMSOS, USP32P2, WFDC10B, WFDC3, WTAPP1, YES1, ZNF286B) и координации сплайсинга 14 генов (ACOT8, CCDC144B, CD40, CTD-2303H24.2, ENOSF1, EVPLL, LGALS9C, MYO15A, SHMT1, SLC12A5, TBC1D28, THOC1, USP32P2, WTAPP1).
Итак, в результате проведённого исследования нами было установлено, что межлокусные взаимодействия 10 изученных SNP генов матриксных металлопротеиназ и генов фолатного цикла материнского организма (rs1799750 [1G>2G] MMP1, rs243865 [C>T] MMP2, rs3025058 [6A>5A] MMP3, rs11568818 [T>C] MMP7, rs17577 [G>A] MMP9, rs1805087 [A>G] MTR, rs1801394 [G>A] MTRR, rs1979277 [C>T] SHMT1, rs699517 [C>T] TYMS, rs2790 [A>G] TYMS) ассоциированы с весом новорожденного. При этом стоит отметить, что наиболее значимый вклад в эти ассоциации вносит полиморфный локус rs1979277 гена серингидроксиметилтрансферазы (SHMT1).
Согласно базам данных по функциональной геномике rs1979277 SHMT1 обладает важными функциональными эффектами в организме. По данным онлайн ресурса HaploReg rs1979277 SHMT1 расположен в области регуляторных мотивов ДНК, где происходит взаимодействие с тремя регуляторными белками, которые регулируют процессы транскрипции: ZEB-1, NKX2 и DMRT2. Разница в показателях логарифма отношения шансов (log-odds) (LOD) для альтернативного аллеля Т и референсного аллеля C составляет для ZEB-1 – 1,2, для NKX2 – -3,0 и для DMRT2– -1,6. Кроме того, с rs1979277 SHMT1 тесно сцеплены (коэффициент корреляции r2≥0,80) 79 SNP, которые располагаются в эволюционно-консервативных областях, сайтах связывания с регуляторными белками (>20 белков), регионе гиперчувствительности к ДНКазе (>50 тканей), в сайтах модификации гистонов, маркирующих промоторы и энхансеры в различных тканях и органах как у взрослого организма, так и у плода (>15 тканей). При этом стоит отметить, что вес-ассоциированные SNP реализуют функциональные эффекты в тканях и органах, которые непосредственно влияют на вес и рост новорожденного, а именно, в скелетных мышцах, подкожной и висцеральной жировой ткани, щитовидной железе и др.
Данный SNP локализован в экзоне гена SHMT1, обуславливая замену лейцина на фениаланин в положении 474 фермента серингидроксиметилтрансферазы 1, что может снижать активность фермента и приводить к снижению уровней фолатов. Также rs1979277 SHMT1 сцеплен с rs8080966 гена SMCR8, приводящим к аминокислотной замене пролина на лейцин в положении 524 белка SMCR8.
Ген SMCR8 (ген-кандидат хромосомного региона синдрома Смита-Магениса, англ. Smith-Magenis syndrome chromosomal region candidate gene 8 protein) кодирует белок, который является субъединицей белкового комплекса SMCR8-C9ORF72. В опытах на мышиных моделях с использованием эмбриональных стволовых клеток Yang et al., показали, что нокаут гена SMCR8 приводил к нарушению индукции процесса аутофагии [42]. Аутофагия, также известная как аутофагоцитоз, представляет собой катаболический механизм, включающий переваривание поврежденных или дисфункциональных клеточных компонентов путем слияния с лизосомами. Кроме того, аутофагия необходима для поддержания клеточного гомеостаза во время пролиферации, дифференцировки и клеточной смерти [43, 44]. Аутофагия также индуцируется в трофобласте на протяжении всего процесса плацентации, что способствует развитию и ремоделированию эндометрия, удалению поврежденных клеточных органелл, а также дифференцировке и инфильтрационной активности трофобласта. Хотя конкретные механизмы этих процессов до конца не изучены, по-видимому, они помогают поддерживать надлежащий материнско-фетальный обмен веществами во время нормального развития плаценты [43]. При этом, как ингибирование, так и усиление аутофагии в ряде исследований было связано со снижением инвазии трофобласта и являлось риском развития преэклампсии [45, 46]. В недавних исследованиях, сравнивающих нормальную беременность с ЗРП, было установлено, что аномальное формирование плаценты, вызывающее ЗРП, связано с дисбалансом клеточного гомеостаза, который усиливает аутофагию в цитотрофобласте [43, 47, 48]. Исходя из роли гена SMCR8 в процессах аутофагии, можно предположить, что rs8080966 SMCR8 может усиливать аутофагию и повышать риск развитии ЗРП.
Полиморфный вариант Т rs1979277 SHMT1 влияет на экспрессию 14 генов (ALKBH5, CCDC144B, CTD-2303H24.2, FAM106A, FAM106A, FOXO3B, LGALS9C, LLGL1, MIEF2, MYO15A, SHMT1, SMCR8, TOP3A, USP32P2) в подкожной и висцеральной жировой ткани, надпочечниках, печени, различных отделах головного мозга, скелетных мышцах, легких, щитовидной железе, крови, культуры клеток фибробластов. 76 SNP из 79 SNP, сцепленных с rs1979277 SHMT1, также определяют изменение транскрипции 28 генов в вышеперечисленных органах и тканях (ALKBH5, CCDC144B, CTD-2303H24.3, DRG2, EPN2-AS1, EVPLL, FAM106A, FLII, FOXO3B, KRT16P1, KRT17P2, LGALS9C, LINC02076, LLGL1, MIEF2, MYO15A, NOS2P2, PRPSAP2, RP11-258F1.1, RP11-258F1.2, RP1-253P7.4, RP1-37N7.1, SHMT1, SMCR8, TOP3A, TVP23B, USP32P2, ZNF286B). Кроме того, установлено, что полиморфный вариант Т rs1979277 SHMT1 связан с более высоким уровнем альтернативного сплайсинга гена CTD-2303H24.2 в скелетных мышцах и щитовидной железе, а также пониженным уровнем альтернативного сплайсинга гена SHMT1 в подкожной и висцеральной жировой ткани, надпочечниках, культуры клеток фибробластов, печени, легких, скелетных мышцах, щитовидной железе, крови, различных отделах головного мозга.
Ген SHMT1 кодирует цитоплазматический фермент серингидроксиметилтрансферазу, являющуюся важным ферментом в метаболическом пути фолатов: она обратимо катализирует превращение серина и тетрагидрофолата в глицин и 5,10-метилентетрагидрофолат (5,10-метилен-ТГФ), соответственно [49]. Эти реакции, с одной стороны, играют ключевую роль в реакциях синтеза пуринов, тимидилата, метилирования ДНК, а с другой стороны служат для превращения гомоцистеина в метионин. На следующем этапе метионин превращается в S-аденозилметионин, выступающий в качестве донора для метилирования дезоксирибонуклеазы, рибонуклеазы и белков [49]. Миссенс-мутация, обусловленная rs1979277 SHMT1, хотя и не влияет на каталитическую активность фермента, но нарушает ядерный транспорт SHMT1 и последующий синтез тимидилата, а также приводит к накоплению измененного фермента SHMT1 в цитоплазме, где он может ингибировать клеточные реакции метилирования, связывая метилен-ТГФ и снижая синтез S-аденозилметионина и способствует накоплению гомоцистеина в крови [50]. В свою очередь гипергомоцистеинемия в материнском организме вызывает дисфункцию эндотелиальных клеток сосудов матери и плаценты, усиливая воспалительную реакцию и окислительный стресс, приводящие к апоптозу и повышенному риску развития ЗРП или рождению ребенка с низкой массой тела [51].
Результаты проведенных ассоциативных исследований демонстрируют связь материнского аллеля Т rs1979277 SHMT1 с повышенным риском рождения детей с низкой массой тела в группе чернокожих женщин и преждевременных родов, как среди белых, так и среди чернокожих беременных [52]. Так как SHMT1 играет важную роль в индукции метилирования генов и синтеза ДНК, большинство публикаций, посвященных генетической вариативности SHMT1, сосредоточены на изучении распространенных злокачественных образований (лейкоз [53, 54], рак молочной железы [55]). В нескольких исследованиях изучалась роль rs1979277 SHMT1 в развитии сердечно-сосудистых заболеваний [56, 57].
Заключение. Таким образом, в результате проведенного исследования установлено, что межлокусные взаимодействия 10 изученных SNP генов матриксных металлопротеиназ и генов фолатного цикла материнского организма (rs1799750 [1G>2G] MMP1, rs243865 [C>T] MMP2, rs3025058 [6A>5A] MMP3, rs11568818 [T>C] MMP7, rs17577 [G>A] MMP9, rs1805087 [A>G] MTR, rs1801394 [G>A] MTRR, rs1979277 [C>T] SHMT1, rs699517 [C>T] TYMS, rs2790 [A>G] TYMS) ассоциированы с весом новорожденного. Наиболее значимый вклад в эти ассоциации вносит полиморфизм rs1979277 SHMT1.
Информация о финансировании
Финансирование данной работы не проводилось.














Список литературы