
Полиморфные варианты генов глутатион S-трансфераз GSTA4 и GSTP1 как маркеры предрасположенности к псориазу и его клинических особенностей
Aннотация
Актуальность: Псориаз является хроническим иммуноопосредованным заболеванием мультифакториальной природы с неуклонно растущей заболеваемостью, достигающей 2-4% в популяции. Несмотря на выявление более 80 локусов предрасположенности, значительная доля наследственного риска остается необъясненной. Ключевая роль в патогенезе псориаза принадлежит окислительному стрессу и воздействию ксенобиотиков окружающей среды, что делает ферменты системы глутатион S-трансфераз, обеспечивающие детоксикацию и антиоксидантную защиту, перспективными кандидатами для изучения. Цель исследования: Провести комплексный анализ вовлеченности полиморфных вариантов генов глутатион S-трансфераз GSTA4 и GSTP1, активно экспрессирующихся в коже, в молекулярные механизмы развития и клинические проявления псориатической болезни. Материалы и методы: В исследование включены 474 пациента с псориазом и 470 здоровых лиц восточнославянского происхождения. Генотипирование 9 однонуклеотидных полиморфизмов генов GSTA4 и GSTP1 выполнено с использованием технологии MALDI-TOF масс-спектрометрии. Проведен анализ ассоциаций с риском развития заболевания, клиническими фенотипами (локализация и характер поражений, псориатический артрит, ониходистрофия) с применением регрессионного анализа и пермутационного теста. Выполнено функциональное аннотирование SNPs с использованием баз eQTL и эпигенетических данных. Результаты: Не выявлено ассоциаций исследуемых полиморфизмов с общим риском псориаза, однако установлена связь варианта rs1790753 GSTP1 с повышенным риском заболевания у женщин (OR=1,31 95%CI 1,01-1,70, Pperm=0,03). Обнаружены многочисленные ассоциации SNPs генов GSTA4 и GSTP1 с поражением различных анатомических зон (верхние и нижние конечности, туловище, ногти), патоморфологическими изменениями (инфильтрация, феномен Кебнера) и развитием псориатического артрита с вовлечением конкретных суставных групп. Функциональное аннотирование подтвердило регуляторный потенциал изученных вариантов и их тканеспецифичное влияние на экспрессию генов в коже. Заключение: Гены GSTA4 и GSTP1 выступают не как факторы общей предрасположенности к псориазу, а как динамические модуляторы его клинического разнообразия, определяющие локализацию поражений, риск артрита и характер течения через регуляцию тканеспецифичной экспрессии генов. Полученные результаты создают основу для разработки персонализированных стратегий прогнозирования, профилактики и терапии псориаза с учетом генетического профиля пациента.
Введение. Современная концепция рассматривает псориаз как хроническое иммуноопосредованное заболевание мультифакториальной природы, в патогенезе которого ключевую роль играет комплексное взаимодействие генетической предрасположенности с различными факторами среды [1, 2]. Это взаимодействие запускает каскад аутовоспалительных и аутоиммунных реакций, приводящих к развитию выраженного системного воспаления, что определяет хроническое и рецидивирующее течение болезни. Псориаз является одним из наиболее часто встречающихся хронических дерматозов. Заболеваемость псориазом в мире оценивается в 2-4% населения планеты [3]. Частота выявления новых случаев увеличилась с 92 (1990 г.) до 99 (2017 г.) на 100 тыс. населения [4] и продолжает неуклонно расти. В Российской Федерации (РФ) также наблюдается рост заболеваемости псориазом, который составил 14% во всех возрастных группах в период с 2010 по 2019 гг. [5].
Несмотря на то, что псориаз является хроническим дерматозом, его клинические проявления не ограничиваются только поражением кожных покровов. Рассматривая псориаз как системное заболевание, многими исследователями употребляется термин «псориатическая болезнь» или «псориатический марш». Данный термин описывает сочетанное поражение кожи, костно-суставной, сердечно-сосудистой, мочевыделительной, эндокринной систем, печени и других органов у пациентов на фоне течения псориаза [6].
Генетические исследования псориаза, включая полногеномные ассоциативные исследования (GWAS), позволили идентифицировать свыше 80 локусов, ассоциированных с предрасположенностью к заболеванию [7, 8, 9]. Исторически первыми и наиболее значимыми были открыты локусы PSORS1 (6p21.3) и PSORS2 (17q25), которые подтвердили ведущую роль наследственности [10, 11, 12]. В дальнейшем были выявлены ключевые гены, участвующие в приобретённом иммунитете (IL23R, IL12B) и барьерной функции кожи (LCE, DEFB) [13-16]. Эти данные позволили перейти от рассмотрения псориаза как дерматоза к пониманию его как системного иммуноопосредованного заболевания с ярко выраженной генетической компонентой. Тем не менее, значительная доля наследственного риска псориаза остаётся необъяснённой – это явление известно как «недостающая наследуемость» (missing heritability) [17]. Данный факт подтверждает сложность мультифакториальной природы псориаза, в развитие которого вносят вклад многочисленные гены слабого эффекта, редкие генетические варианты, а также их взаимодействия между собой и с факторами окружающей среды.
Несмотря на значительный прогресс, патогенез псориаза остается не до конца расшифрованным: ключевые механизмы, определяющие инициацию болезни, её клиническую гетерогенность и связь с системными осложнениями, требуют дальнейшего изучения. Известно, что при псориазе наблюдаются патологические изменения в системе редокс-гомеостаза, проявляющиеся ослаблением антиоксидантной системы и повышенным образованием свободных радикалов, что в конечном итоге приводит к окислительному стрессу [18, 19] – состоянию дисбаланса между окислителями и антиоксидантами в пользу первых, вызывающему нарушение окислительно-восстановительной сигнализации и контроля и/или молекулярные повреждения. Окислительные повреждения стимулируют гиперпролиферацию кератиноцитов и нарушения микроциркуляции [20, 21]. Кроме того, обширные литературные данные свидетельствуют о том, что маркеры окислительного стресса повышены при псориазе и связаны с длительностью и тяжестью заболевания [22].
Недавними эколого-эпидемиологическими исследованиями установлено, что ксенобиотики окружающей среды являются важной детерминантой развития и прогреccирования псориаза [23-26]. В частности, показано, что воздействие смесей перхлоратов, нитратов и тиоцианатов, а также этилпарабена связано с повышенной распространённостью псориаза [27]. Загрязнение воздуха значительно увеличивает вероятность развития псориаза, и величина этого повышения сопоставима с риском, связанным с воздействием табачного дыма, при этом установлено, что кожа является недостаточным барьером для проникновения смешанных ксенобиотиков в организм [28]. Недавнее крупномасштабное проспективное когортное исследование 451 064 участников из Британского биобанка показало, что длительное воздействие различных загрязнителей воздуха положительно коррелирует с повышенным риском развития псориаза, особенно у лиц с высоким генетическим риском этого заболевания [25]. Таким образом, полученные данные позволяют рассматривать химические контаминанты окружающей среды в качестве значимого этиологического компонента в развитии псориаза и обосновывают целесообразность дальнейших исследований молекулярных механизмов болезни в контексте токсикогенетической концепции мультифакториальной патологии [29]. Есть все основания полагать, что химический стресс и связанный с ним окислительный стресс [30, 31], вызванные попаданием в организм человека различных молекул химических веществ и их соединений, наряду с механической травмой кожи и психоэмоциональным стрессом, могут играть ключевую роль в манифестации и прогрессировании псориаза.
В системе биотрансформации ксенобиотиков, где ключевая детоксикационная роль принадлежит реакциям второй фазы (конъюгации), особое место занимают ферменты семейства глутатион S-трансфераз (GST), выступающие центральным звеном в обезвреживании широкого спектра электрофильных соединений [32, 33, 34]. Учитывая наличие функциональных неравнозначных аллельных вариантов в структуре GST, данное семейство генов стало привлекательным объектом для генетико-ассоциативных исследований различных заболеваний [35-38]. В контексте данного вопроса ферменты метаболизма глутатион S-трансфераз, участвующих в нейтрализации ксенобиотиков в кожных покровах, могут выступать как один из основных патогенетических путей развития псориаза. Функциональное состояние ферментов глутатион S-трансфераз и уровень их экспрессии в коже, которые также обусловлены наличием ДНК-полиморфизмов в структуре соответствующих генов, при воздействии различных эндогенных и экзогенных факторов, потенциально могут иметь ключевое значение в патогенезе данного заболевания. Несмотря на проведение крупных международных полногеномных исследований, выявивших десятки ассоциированных с псориазом полиморфизмов, многие молекулярные аспекты его патогенеза, в частности роль системы детоксикации ксенобиотиков, остаются недостаточно изученными. В литературе имеются единичные данные о связи полиморфных вариантов генов глутатион S-трансфераз с риском развития и клиническими проявлениями псориаза [39, 40, 41]. При этом проведённые исследования были преимущественно сфокусированы на генах глутатион S-трансфераз классов theta (GSTT1) и mu (GSTM1). В нашем недавнем исследовании нами установлены ассоциации псориаза с полиморфными вариантами гена GSTO1 [42]. Исследований, направленных на анализ связи полиморфизма генов глутатион S-трансфераз GSTA4 и GSTP1, активно экспрессирующихся в коже, с развитием и клиническими особенностями псориаза до настоящего времени не проводилось ни в России, ни за рубежом. Всё вышеизложенное указывает на необходимость проведения комплексного анализа вовлечённости полиморфных вариантов генов глутатион S-трансфераз в этиопатогенез и клинические проявления псориаза.
Цель исследования. Провести комплексный анализ вовлеченности полиморфных вариантов генов глутатион S-трансфераз GSTA4 и GSTP1 в молекулярные механизмы развития и клинические проявления псориатической болезни.
Материалы и методы исследования. Для проведения данного исследования были использованы образцы ДНК, клинические и анкетные данные 944 неродственных индивидуумов, включая 474 пациента с диагнозом «псориаз» и 470 условно здоровых лиц (группа контроля). Все пациенты, включённые в исследовательскую группу, имели восточнославянское происхождение (преимущественно русские, а также украинцы, белорусы и потомки от смешанных браков между представителями данных национальностей) и являлись уроженцами преимущественно Курской области. Этническая принадлежность определялась на основании анкетирования по критерию происхождения пробанда: наличие у него родителей, а также бабушек и дедушек по обеим линиям, относящихся к восточнославянским национальностям. Основная группа пациентов с псориазом была сформирована сплошным методом в период с 2018 по 2021 год на клинической базе следующих медицинских учреждений: ОБУЗ «Медвенская центральная районная больница» (посёлок Медвенка Курской области), ООО «Центр медицинских осмотров и профилактики» (г. Курск) и Кожно-венерологического диспансера Курской областной клинической многопрофильной больницы (г. Курск). Контрольная группа была сформирована на основе материалов биобанка Научно-исследовательского института генетической и молекулярной эпидемиологии Курского государственного медицинского университета, собранных в рамках ранее проведённых генетико-эпидемиологических исследований различных мультифакториальных заболеваний. Все участники исследования предоставили письменное информированное добровольное согласие. Протокол исследования был одобрен локальным этическим комитетом при Курском государственном медицинском университете (протокол № 4 от 09.04.2018).
Критериями включения пациентов в основную группу являлись: верифицированный врачом-дерматологом диагноз «псориаз»; восточнославянское происхождение, определяемое генеалогическим методом; отсутствие иных хронических заболеваний, манифестировавших до появления первых клинических признаков псориаза; предоставление письменного информированного добровольного согласия на участие. В критерии исключения вошли: не восточнославянское происхождение; отказ от участия; наличие беременности на момент включения в исследование; наличие тяжёлых системных заболеваний в стадии обострения (включая ревматологические заболевания и злокачественные новообразования), манифестировавших до дебюта псориаза; хронические инфекционные заболевания (вирусные гепатиты, ВИЧ-инфекция, туберкулёз лёгких); а также проведение пациенту в течение месяца, предшествующего включению, местной терапии псориаза либо системной терапии (включая фото- и химиотерапию) по поводу любого заболевания в течение последних трёх месяцев. Критериями включения в контрольную группу служили отсутствие хронических заболеваний, восточнославянское происхождение и предоставление информированного согласия. Основаниями для исключения являлись не восточнославянское происхождение и отказ от участия.
Диагноз у пациентов основной группы устанавливался на основании типичной клинической картины псориаза: наличие эпидермально-дермальных папул и бляшек любой локализации с шелушением мелкими серебристо – белыми чешуйками на поверхности, наличие подногтевых псориатических элементов в виде пятна грязно – желтого цвета (симптом «масляного пятна») и множественных точечных углублений на поверхности ногтей (симптом «наперстка») [43]. В исследование были включены пациенты с классическим бляшечным псориазом, ладонно-подошвенным, себорейным и псориазом волосистой части головы; генерализованным пустулезный псориазом Цумбуша, инверсным псориазом; каплевидным псориазом; и псориатической эритродермией, а также пациенты с сопутствующими поражениями костно-суставной системы (псориатический артрит) и ногтевого аппарата (псориатическая ониходистрофия). Клиническая оценка псориаза проводилась с использованием индекса площади и тяжести течения псориаза – “Psoriasis Area and Severity Index” или PASI [44]. Для оценки тяжести течения заболевания использовался эпидемиологический скрининговый тест на выявление псориатического артрита – “Psoriasis Epidemiology Screening Test”, или “PEST” [45]. Для клинической оценки псоритаической ониходистрофии был использован индекс площади и тяжести течения псориаза ногтей (Nail Psoriasis Area and Severity Index или NAPSI) [46]. Шкала стрессочувствительности Холмса-Рея [47] и дерматологический индекс оценки качества жизни – Dermatological Quality of Life Index или DLQI [48] использовались для оценки психологического состояния пациентов и их качества жизни, соответственно. Для оценки общего соматического статуса использовали индекс массы тела (ИМТ или BMI) [49].
Все участники исследования прошли анкетирование, которое проводил врач-дерматолог с использованием специально разработанной анкеты. Анкета содержала следующие блоки: 1) анализ факторов риска (употребление алкоголя, курение, наличие психоэмоционального стресса, контакт с агрессивными химическими веществами, особенности питания); 2) клинический и семейный анамнез (возраст и характер манифестации заболевания, его сезонность, частота и характер обострений, наличие псориаза и других хронических патологий у родственников); 3) объективные клинические данные (площадь и характер псориатических поражений, наличие коморбидных состояний, анамнез ранее проводимой терапии). Данные анкетирования заносились в электронную базу MS Excel и кодировались в соответствии со шкалой измерения переменных: в виде дихотомических (0 – отсутствие признака, 1 – наличие признака), порядковых (три и более градаций) или количественных фенотипических признаков.
Для отбора генов, кодирующих ферменты глутатион S-трансферазы (GST), с высокой базовой экспрессией в ткани кожи, была использована база данных GTEx Portal (https://gtexportal.org). В результате на основе анализа транскрипционного профиля для исследования были выбраны гены GSTA4 (6p12.2) и GSTP1 (11q13.2). Критерием отбора генов послужил высокий уровень их экспрессии в коже нижних конечностей (транскриптомная база “Skin-Sun Exposed/Lower leg”) и/или нижней части живота (транскриптомная база “Skin-Not Sun Exposed/Suprapubic”). Анализ и отбор однонуклеотидных полиморфизмов (single nucleotide polymorphism, SNP) генов глутатион S-трансфераз проводился с учётом гаплотипической структуры генов, функциональных свойств SNP и частот минорных аллелей с использованием пакета биоинформатических инструментов SNPinfo Web Server [50]. Для обеспечения максимального покрытия генов представляли интерес в первую очередь таггерные SNP (tagSNP) — полиморфные маркеры, репрезентирующие группы локусов (гаплотипы), находящихся в сильном (D' ≥ 0.8) неравновесии по сцеплению [51]. Отбор tagSNP осуществляли с помощью биоинформатического инструмента GenePipe (https://snpinfo.niehs.nih.gov/snpinfo/selegene.html) на основе референсной гаплотипической структуры европеоидной популяции из проекта HapMap. Оценку функциональной значимости SNP выполняли с использованием алгоритма FuncPred платформы SNPinfo (https://snpinfo.niehs.nih.gov/snpinfo/snpfunc.html). Отбор однонуклеотидных полиморфизмов (SNP) проводился на основании четырёх ключевых критериев: (1) частота минорного аллеля (MAF) не менее 1% в европейских популяциях по данным базы Ensembl; (2) принадлежность SNP к локусам количественных признаков, регулирующим экспрессию (eQTL) и/или сплайсинг (sQTL), в ткани кожи (участков, подверженных и/или не подверженных инсоляции — нижние конечности и нижняя часть живота соответственно) согласно ресурсу GTEx; (3) уровень неравновесия по сцеплению (r²) между отобранными маркерами менее 0,8 в европейской популяции проекта HapMap; (4) техническая возможность их совместного генотипирования в рамках одной мультиплексной панели методом MALDI-TOF масс-спектрометрии. В результате было отобрано 9 SNPs генов глутатион S-трансфераз: 5 SNPs (rs7496, rs17614871, rs12524274, rs316133, rs2274760) гена GSTA4 и 4 SNPs (rs1790753, rs1695, rs749174, rs1138272) гена GSTP1.
Выделение геномной ДНК проводили методом фенольно-хлороформной экстракции с последующей преципитацией этанолом. Все молекулярно-генетические этапы работы — от выделения ДНК до генотипирования – были выполнены в НИИ генетической и молекулярной эпидемиологии КГМУ в период с 2021 по 2022 год. Детекция однонуклеотидных полиморфизмов в генах глутатион S-трансфераз проводилась с использованием технологии iPLEX, позволяющей осуществлять мультиплексное генотипирование на масс-спектрометрическом анализаторе MALDI-TOF MassARRAY 4 компании Agena Bioscience (США) согласно протоколу, описанному нами ранее [52]. Перед масс-спектрометрическим анализом образцы ДНК были амплифицированы с помощью мультиплексной полимеразной цепной реакции (ПЦР), нацеленной на исследуемые участки генов GST. Реакцию проводили на амплификаторе CFX-96 (BioRad, США). Дизайн праймеров и формирование ПЦР-мультиплекса для последующего генотипирования SNP выполняли с использованием пакета MassARRAY Assay Design Suite компании Agena Bioscience. Синтез олигонуклеотидных праймеров для iPLEX-генотипирования осуществила компания «Евроген» (Москва, http://evrogen.ru). Для контроля качества генотипирования методом случайной выборки были отобраны 95 образцов, которые повторно генотипировали по всем исследуемым локусам на анализаторе MALDI-TOF MassARRAY 4. Результаты повторного анализа полностью соответствовали первичным данным в 99% случаев, что подтверждает высокую воспроизводимость метода и качество проведённого генотипирования.
Для сравнения групп по категориальным переменным использовали критерий χ², а по количественным переменным – t-критерий Стьюдента (для показателей с нормальным распределением) или U-критерий Манна – Уитни (для показателей, распределение которых отличалось от нормального). Соответствие распределения генотипов равновесию Харди – Вайнберга оценивали с помощью точного критерия Фишера. Анализ ассоциации аллелей и генотипов с риском развития псориаза проводился путём расчёта отношения шансов (Odds Ratio, OR) с поправкой на пол и возраст. Данный показатель отражает, во сколько раз вероятность наличия заболевания выше у носителя определённого аллеля или генотипа по сравнению с лицами, его не имеющими [53]. Отношения шансов (OR) с 95% доверительными интервалами (95% CI), отражающие ассоциацию SNP с псориазом, рассчитывали в рамках аддитивной генетической модели с использованием критерия линейного тренда Кохрана-Армитиджа для таблиц сопряжённости 2×3 в программе PLINK v1.9 [54]. Влияние SNP на количественные фенотипы (площадь поражения, индекс PASI, возраст дебюта заболевания и частоту обострений) в группе пациентов с псориазом оценивали методом линейного регрессионного анализа также в программе PLINK v1.9. Для оценки уровня неравновесия по сцеплению между локусами, а также для анализа ассоциаций гаплотипов генов GST с риском развития псориаза использовали программу Haploview версии 4.2 [55]. Статистическую значимость ассоциаций (Pperm) оценивали с помощью адаптивного пермутационного теста. Функциональное аннотирование полиморфных вариантов генов и оценка их регуляторного потенциала осуществлялась с использованием различных биоинформатических инструментов: GTEx portal (https://gtexportal.org), eQTLGen Consortium (www.eqtlgen.org), VannoPortal (http://www.mulinlab.org/vportal/index.html) и Haploreg 4.2 (https://pubs.broadinstitute.org/mammals/haploreg/haploreg.php).
Результаты исследования. Средний возраст пациентов с псориазом (44,3 года) был на 11 лет ниже, чем в контрольной группе (P<0,001). Поскольку псориаз является заболеванием с пожизненным риском, включение в контрольную группу лиц старшего возраста повышает достоверность вывода об отсутствии у них заболевания на момент исследования. Минимальный возраст в когорте пациентов составил 8 лет, максимальный – 84 года. Группы пациентов с псориазом и контроля не отличались по полу (P>0,05). Наиболее частыми локализациями псориатических поражений были: верхние (80%) и нижние (57%) конечности, а также голова (48%). Средний возраст манифестации (дебюта) псориаза в исследуемой группе составил 30 лет, а средняя продолжительность заболевания – 14 лет. Псориатический артрит (ПсА) любой локализации был диагностирован у 27% пациентов (n=128). Среди суставных групп наиболее часто поражались коленные суставы (12,4%) и суставы пальцев кисти (12,7%). Локтевые и кистевые суставы были вовлечены у 7% пациентов каждый. Реже наблюдалось поражение голеностопных (5,1%), тазобедренных (4,4%), суставов пальцев стопы (4,9%) и большого пальца кисти (3,8%). Наименьшая частота поражения отмечена для плечевых суставов (2,3%). Таким образом, в исследуемой когорте преобладал периферический вариант ПсА с частым вовлечением суставов пальцев кисти и коленных суставов. Псориатическая ониходистрофия была диагностирована у 59,3% пациентов. Сочетанное поражение суставов и ногтевых пластин наблюдалось у 32% больных.
Частоты генотипов исследуемых полиморфных вариантов генов ферментов глутатион S-трансфераз в исследованной нами популяции находились в равновесии Харди-Вайнберга (P>0,05). Один из прогенотипированных SNP rs17614871 в гене GSTA4 оказался трех-аллельным (аллели T,A,G). Частоты генотипов данного локуса были следующими: T/T 54,7%, T/A 29,1%, G/T 10,6%, A/A 3,3%, G/A 2,1% и G/G 0,2%. Из-за методологических ограничений статистической обработки данный полиморфный вариант был исключен из исследования. Сопоставление частот аллелей GST с другими европейскими популяциями с использованием интернет базы данных Ensembl (https://www.ensembl.org/index.html) показало, что, в целом, частоты аллелей изучаемых полиморфных вариантов генов существенно не отличаются от других европеоидных популяций (финнов, британцев, испанцев, итальянцев и резидентов штата Юта США - выходцев из Северной и Западной Европы), которые являются частью международного проекта HapMap третьей фазы. Различие в частоте минорного аллеля в сравнении с объединенной европеоидной выборкой выявлено для полиморфизма rs7496 гена GSTA4 (MAF=0,096 P=0,02).
Проведено сравнение частот аллелей и генотипов полиморфных вариантов генов глутатион S-трансфераз между группами больных псориазом и контроля. Результаты анализа ассоциации аллелей и генотипов полиморфных вариантов генов, кодирующих ферменты GST, с риском развития псориатической болезни в общей выборке пациентов представлены в таблицах 1 и 2, соответственно.
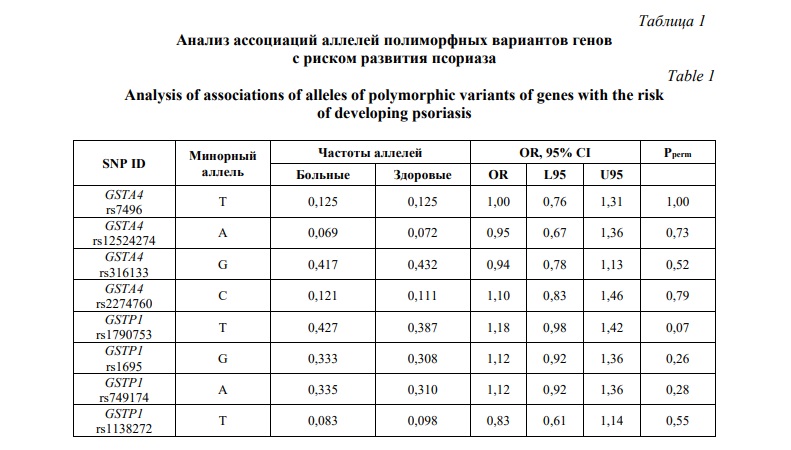
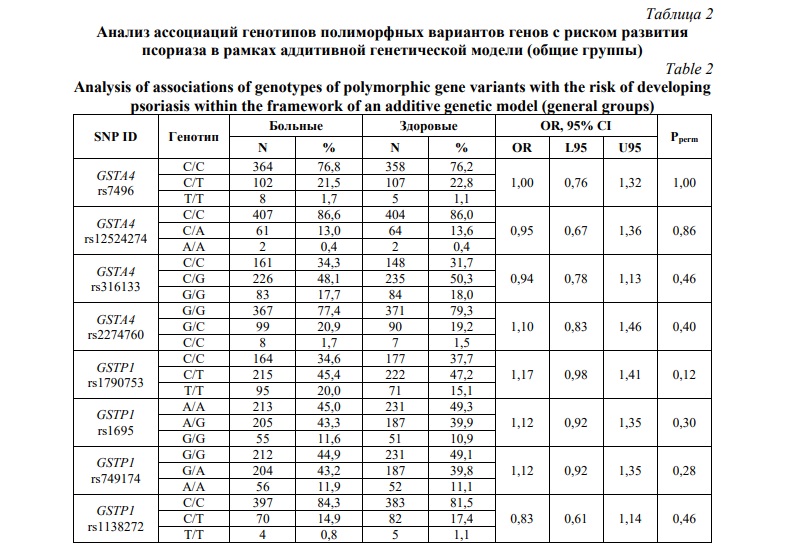
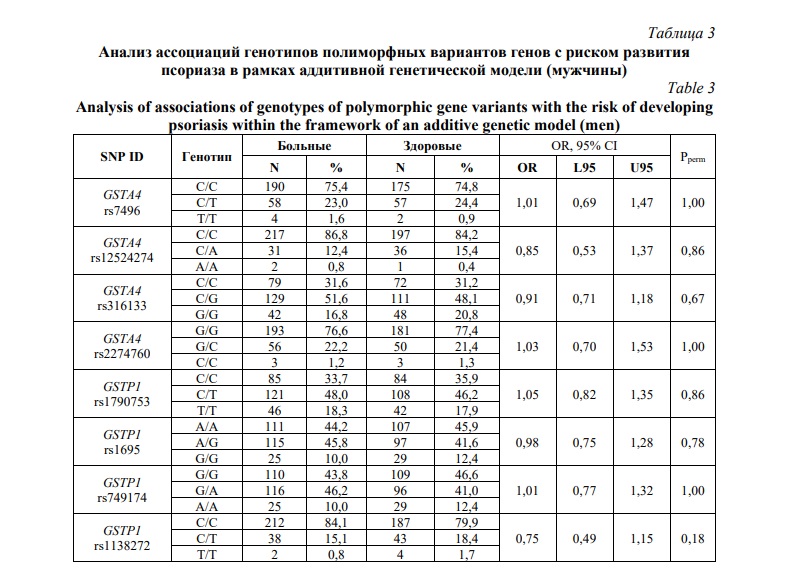
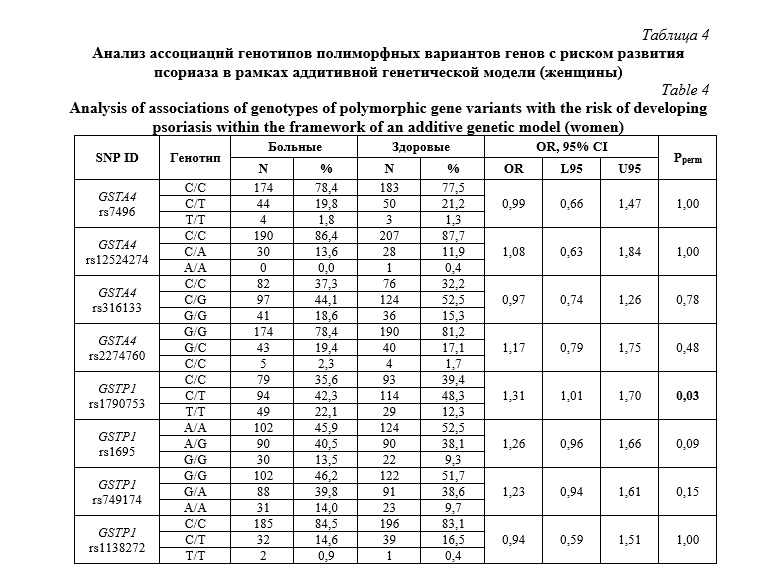
Как видно из таблицы 1, среди проанализированных полиморфных вариантов генов только аллель rs1790753-T гена GSTP1 имел отчетливую тенденцию к ассоциации с повышенным риском развития болезни, но уровень статистический значимости (Pperm=0,07) не достигал принятого в исследовании порогового значения. Не было установлено статистически значимых ассоциаций аллелей и генотипов гена GSTA4 с риском развития псориаза. Учитывая возможность полового диморфизма в ассоциациях ДНК-маркеров с предрасположенностью к мультифакториальным заболеваниям, в том числе к псориазу [56, 57], представлял интерес анализ ассоциаций аллелей и генотипов GST с развитием псориатической болезни раздельно у мужчин и женщин. У женщин выявлена ассоциация с псориазом в отношении минорного аллеля rs1790753-T гена GSTP1 (OR=1,33 95%CI 1,02-1,73, Pperm=0,04). В таблицах 3 и 4, представлены результаты анализа ассоциаций генотипов GSTA4 и GSTP1 с развитием псориаза у мужчин и женщин, соответственно. Как можно увидеть из таблицы 3, не было выявлено статистически значимых ассоциаций исследуемых генов GST с риском развития псориаза у мужчин (Pperm>0,05). Однако (таблица 4), полиморфизм rs1790753 гена GSTP1 ассоциировался с повышенным риском развития псориатической болезни у женщин в рамках как аддитивной генетической модели (OR=1,31 95%CI 1,01-1,70, Pperm=0,03). Генотипы, включающие минорный аллель T (rs1790753-C/T и rs1790753-T/T), более чем в 2 раза увеличивали шансы развития псориатической болезни у женщин (OR=2,02 95%CI 1,22-3,34, Pperm=0,006). В отношении других полиморфных вариантов генов GST статистически значимых ассоциаций с риском развития псориаза не установлено, как у женщин, так и у мужчин.
Анализ гаплотипов, в отличие от изолированного изучения отдельных полиморфизмов, позволяет оценить кумулятивный эффект конаследуемых аллелей в пределах одного гена или генного кластера. Такой подход является методологической основой для более целостного понимания вклада сложных генетических структур в предрасположенность к мультифакториальным заболеваниям [14]. В рамках настоящего исследования был проведён анализ ассоциаций гаплотипов генов глутатион S-трансфераз GSTA4 и GSTP1 с риском развития псориаза, направленный на выявление специфических комбинаций аллелей, которые могут определять устойчивость или восприимчивость к заболеванию. Анализ ассоциаций гаплотипов с псориазом был проведен, как в общих группах, так и в группах, стратифицированных по полу.
В таблице 5 представлены гаплотипы гена GSTA4 результаты анализа их ассоциации c риском развития псориатической болезни. Установлено 5 гаплотипов гена GSTA4: H1 (C-C-C-G) – самый распространенный с частотой 55-57%, H2 (T-C-G-G), H3 (C-C-G-C) иH4 (C-C-G-G) с частотой порядка 10-13%, а также самый редкий (частота 7-8%) гаплотип H2 (C-A-G-G). Сравнительный анализ частот гаплотипов между группами больных псориазом и здоровых индивидов не выявил статистически значимых различий в их распределении (Pperm>0,05).
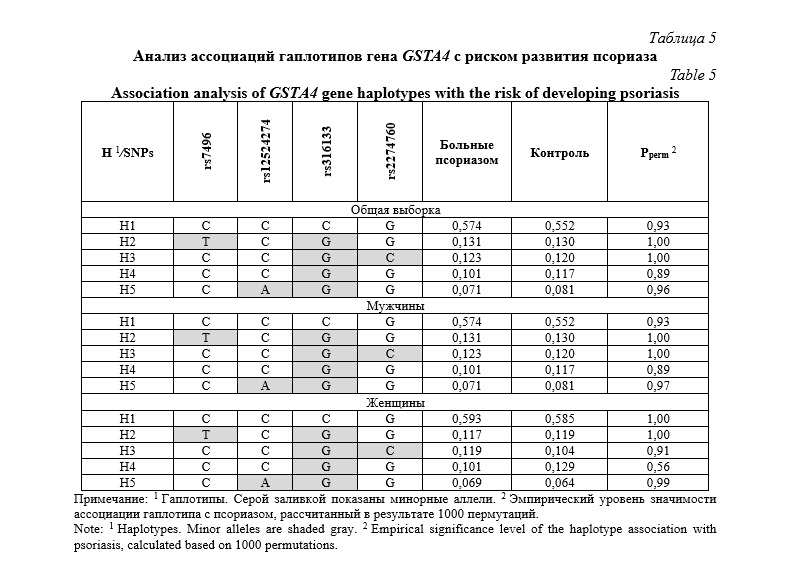
В таблице 6 представлены частоты гаплотипов гена GSTP1 и результаты их сопоставления между группами больных псориазом и контроля. Было выявлено пять частых гаплотипов гена GSTP1 с частотой встречаемости более 5%. Среди них гаплотип H1 (C-C-C-G) являлся преобладающим, составляя более 50% от общего числа. Сравнительный анализ частот гаплотипов гена GSTP1 не выявил статистически значимых различий между группами пациентов с псориатической болезнью и контроля, как при сопоставлении в общей группе пациентов, так и среди мужчин. Однако, у женщин самый частый гаплотип H1 был ассоциирован с пониженным риском развития псориаза (OR=0,71 95%CI 0,49-1,03, Pperm=0,015). Полиморфизм rs1790753 гена GSTP1, ассоциированный с повышенным риском псориаза у женщин, позитивно коррелировал с SNP rs1695 (D=0.1276, D¢=0.6713, P=2,0×10-16), rs749174 (D=0,1279, D¢=0,6696, P=2,0×10-16) и rs1138272 (D=0,0528, D¢=0,9863, P=2,0×10-16).
Учитывая ключевую функцию глутатион S-трансфераз в детоксикации ксенобиотиков, нейтрализации окислительного стресса и регуляции воспаления – процессов, центральных в патогенезе псориаза, – анализ ассоциаций полиморфизмов GST с особенностями кожного поражения (локализацией, степенью тяжести, распространённостью бляшек) и внекожными проявлениями (в первую очередь псориатическим артритом и другими коморбидностями) позволяет глубже понять механизмы фенотипической гетерогенности заболевания. В этой связи нами проведен анализ ассоциаций полиморфных вариантов генов GSTA4 и GSTP1 с клинически особенностями псориаза. Первым этапом анализа была оценка связи полиморфизма генов GST с распространенностью и характером поражения различных участков тела с псориазом методом регрессионного анализа с поправкой на пол пациентов.
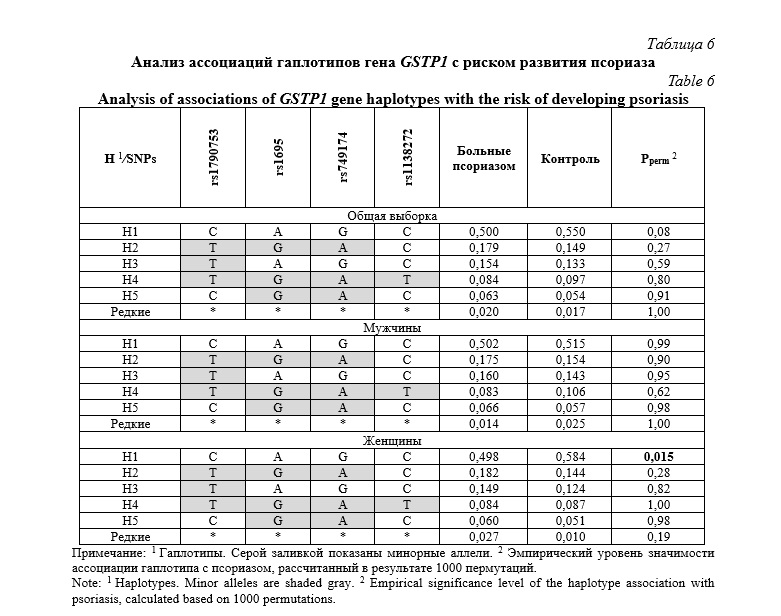
В таблице 7 представлены сводные данные по статистически значимым ассоциациям исследуемых локусов с поражением различных участков тела у пациентов с псориазом. Анализ показал, что с поражением верхних конечностей были ассоциированы полиморфные варианты rs316133 гена GSTA4 (β=0,501, Pperm=0,05) и rs749174 гена GSTP1 (β=-0,530, Pperm=0,03). Полиморфизм rs1790753 гена GSTP1 (β=-0,391, Pperm=0,036) был связан с псориатическим поражением на туловище. SNP rs1790753 гена GSTP1 также ассоциировался с поражением кожи поясничной области (β=-1,804, Pperm=0,03). С поражением ногтей были связаны полиморфные варианты rs1695 (β=-0,492, Pperm=0,016) и rs749174 (β=-0,522, Pperm=0,015) гена GSTP1.
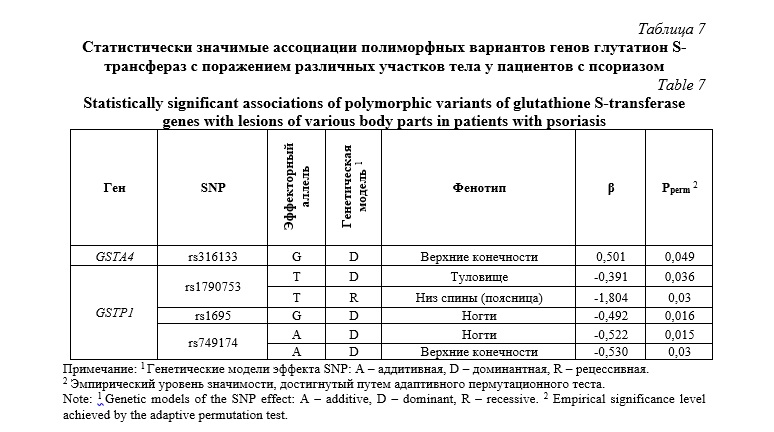
В таблице 8 представлены сводные данные по статистически значимым ассоциациям полиморфных вариантов генов GST с патоморфологическими изменениями кожи при псориазе. Хотя нами не было установлено статистически значимых ассоциаций исследуемых локусом с индексом PASI – показателем тяжести и распространенности псориаза, тем не менее, установлен ряд ассоциаций полиморфизмов генов GST с характером распространенности поражений кожи, учитываемых при расчете данного индекса. В частности, установлена статистически значимая ассоциация с площадью псориатического поражения кожных покровов нижних конечностей и rs1695 гена GSTP1 (β=0,189, Pperm=0,037). Полиморфные варианты rs1695, rs749174 и rs1138272 гена GSTP1 также ассоциировались с феноменом Кебнера (Табл. 8).
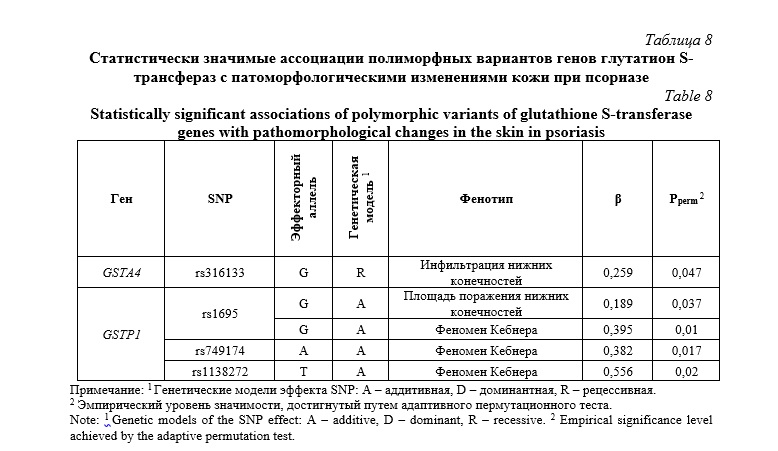
Однако значительная доля выявленных ассоциаций охватывала гистопатоморфологические изменения кожных покровов у пациентов с псориазом. Ключевым компонентом псориатической бляшки является инфильтрация – уплотнение кожи, возникающее из-за комбинации двух основных процессов: аномально высокой пролиферации кератиноцитов и миграции в очаг воспалительных клеток (лимфоцитов и макрофагов) [58]. Мы обнаружили, что с развитием инфильтрации кожных покровов нижних конечностей ассоциирован SNP rs316133 гена GSTA4 (β=0,259, Pperm=0,047).
Затем мы проанализировали связь между полиморфизмами генов GST и риском развития псориатического артрита — наиболее распространенного внекожного осложнения псориаза [59]. В таблице 9 представлены результаты по обнаруженным статистически значимым ассоциациям полиморфных вариантов генов глутатион S-трансфераз с поражением различных суставов у пациентов с псориазом.
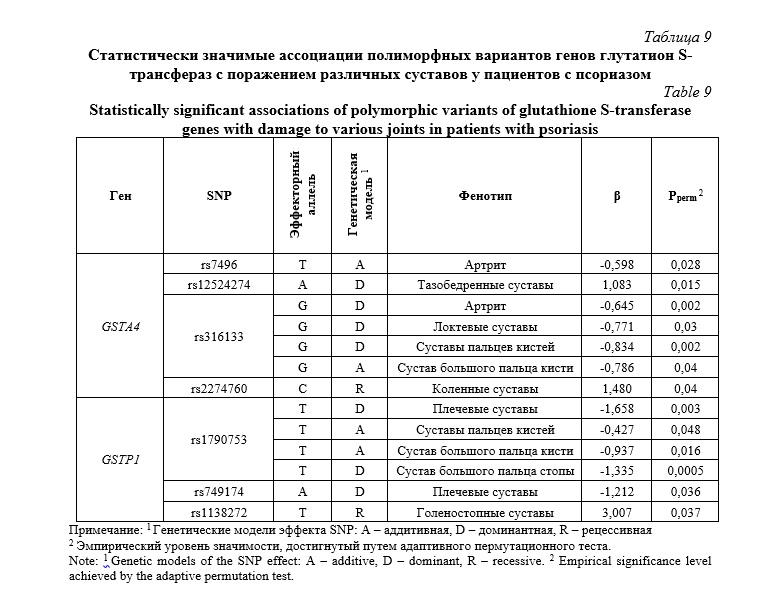
Развитие артрита (независимо от локализации суставного поражения) ассоциировалось с полиморфизмами rs7496 (GSTA4, β=-0,598, Pperm=0,028) и rs316133 (GSTA4, β=-0,645, Pperm=0,002). Также установлены ассоциации определенных полиморфных локусов с преимущественным поражением определенных суставов. Часть изученных полиморфизмов продемонстрировала ассоциацию не с артритом в целом, а с поражением конкретных групп суставов, что указывает на возможную генетическую детерминацию паттерна вовлечения опорно-двигательного аппарата. В первую очередь представляет интерес данные по гено-фенотипическим корреляциям, обнаруженным в отношении поражения крупных суставов. Так, с псориатическим поражением плечевых суставов были ассоциированы полиморфные варианты rs1790753 (β=-1,658, Pperm=0,003) и rs749174 (β=-1,212, Pperm=0,036) гена GSTP1. С псориатическим артритом локтевых суставов был ассоциирован rs316133 гена GSTA4 (β=-0,771, Pperm=0,03). Полиморфизм rs12524274 гена GSTA4 (β=1,083, Pperm=0,015) был ассоциирован с поражением тазобедренных суставов. С поражением коленных суставов был связан полиморфизм rs2274760 гена GSTA4 (β=1,480, Pperm=0,04). SNP rs1138272 гена GSTP1 (β=3,007, Pperm=0,037) коррелировал с артритом голеностопных суставов. Полиморфные варианты генов GST также ассоциировались с поражением мелких суставов верхних и нижних конечностей. Так, полиморфизм rs316133 гена GSTA4 (β=-0,834, Pperm=0,002) и rs1790753 гена GSTP1 (β=-0,427, Pperm=0,048) были связаны с артритом суставов пальцев кистей. Кроме того, SNP rs316133 гена GSTA4 (β=-0,786, Pperm=0,04) и rs1790753 гена GSTP1 (β=-0,937, Pperm=0,016) ассоциировались с поражением сустава большого пальца кисти. Полиморфизм rs1790753 гена GSTP1 (β=-1,335, Pperm=0,0005) также был ассоциирован с артритом большого пальца стопы.
Нами также установлены ассоциации полиморфизма генов GST полиморфных вариантов генов глутатион S-трансфераз с клиническим течением псориаза. Так, со степенью распространенности кожных поражений при первичной клинической манифестации псориаза был ассоциирован полиморфизм rs2274760 гена GSTA4 (β=0,168, Pperm=0,036). Полиморфизм rs12524274 гена GSTA4 (β=0,367, Pperm=0,02) был ассоциирован с частотой рецидивов псориаза. Дерматологический индекс качества жизни пациентов с псориазом коррелировал с rs7496 гена GSTA4 (β=-0,205, Pperm=0,01). Статистически значимых ассоциаций полиморфных вариантов генов GST ни с возрастом манифестации псориаза, ни с показателями стрессоустойчивости (оцененными по тесту социальной адаптации) выявлено не было.
Проведено функциональное аннотирование полиморфных вариантов генов GST с использованием интернет-ресурсов eQTLGen Consortium и GTEx portal. Анализ полиморфизмов гена GSTA4 выявил преимущественно негативные регуляторные эффекты в крови с тканеспецифичными вариациями. Аллели A варианта rs12524274 и C варианта rs2274760 являются значимыми негативными eQTL для собственного гена GSTA4 в крови (Z=-8,61 и Z=-4,49 соответственно), причём rs2274760 в фибробластах кожи проявляет противоположный, позитивный эффект (β=0,12). Наиболее сильным негативным регулятором GSTA4 в крови является аллель G варианта rs316133 (Z=-15,37). Вариант rs7496 (T), не показавший значимого eQTL в крови, специфично повышает экспрессию GSTA4 в коже (β=0,11). Затем были проанализированы эпигенетические маркеры, ассоциированные с полиморфными вариантами гена GSTA4. Для варианта rs12524274 значимых эпигенетических ассоциаций выявлено не было. В доступных данных Roadmap Epigenomics для полиморфизма rs12524274 гена GSTA4 также не описаны мотивы связывания транскрипционных факторов. Биоинформатический анализ для полиморфизма rs2274760 позволил предсказать в его локусе потенциальный сайт связывания для транскрипционного фактора Cdx. Так, полиморфизм rs7496 в гене GSTA4 расположен в участке, способном связывать транскрипционные факторы Hmbox1 и Nanog. Хотя для полиморфизма rs316133 гена GSTA4 не обнаружено характерных эпигенетических модификаций, его локализация в области связывания для транскрипционного фактора AP-1 свидетельствует о возможном прямом регуляторном эффекте.
Анализ eQTL для GSTP1 показал, что основные исследуемые SNP являются мощными регуляторами не собственного гена, а соседних локусов, и находятся в сильном сцеплении. Аллель T варианта rs1790753 — негативный eQTL для GSTP1 в крови (Z=-4,91), но при этом сильнейший негативный регулятор гена DOC2GP (β до -0,35) и позитивный регулятор NDUFV1. Варианты rs1695 (G) и rs749174 (A) демонстрируют идентичные и экстремально сильные eQTL-эффекты (например, для DOC2GP:Z=49,36, β=-0,50), что указывает на маркирование общего функционального гаплотипа. Данные по eQTL для собственного гена GSTP1 для rs1138272 и rs1695 в базах данных отсутствуют. Полиморфизм rs1790753 GSTP1 характеризуется сложной и клеточно-специфичной регуляторной архитектурой в различных типах клеток кожи. В первичных фибробластах кожи он ассоциирован с сигнатурами активной транскрипции (3_TxFlnk), усиленными (12_TxEnhW) и активными (H3K27ac_Enh) энхансерами, а также активными промоторами (H3K4me3_Pro). В первичных кератиноцитах кожи его эпигенетический профиль ограничен активными промоторами (H3K4me3_Pro), тогда как в первичных меланоцитах он вновь демонстрирует признаки активной энхансерной (6_EnhG, 11_TxEnh3, H3K27ac_Enh) и промоторной (H3K4me3_Pro) активности. SNP rs1790753 расположен в участке, предсказанном для связывания транскрипционных факторов ATF3, ELF1, ERalpha-a, NF-kappaB и Zfx. SNP rs749174 демонстрирует консервативный паттерн регуляторной активности во всех основных типах клеток кожи (фибробластах, кератиноцитах, меланоцитах), ассоциируясь с сигнатурами активной транскрипции (3_TxFlnk), регуляторных регионов (9_TxReg), а также активных энхансеров (H3K4me1, H3K27ac) и промоторов (H3K4me3). Дополнительная ассоциация с открытым хроматином (DNase) в эпидермальных кератиноцитах подтверждает его расположение в ключевом доступном регуляторном элементе. Полиморфизм rs749174 расположен в участке, предсказанном для связывания транскрипционных факторов AP-2rep (репрессор на основе AP-2) и RXRA (ретиноид X рецептор альфа). SNP rs1695 демонстрирует консервативный профиль активного регуляторного элемента во всех основных типах клеток кожи, ассоциируясь с сигнатурами транскрипции (3_TxFlnk), регуляторных областей (9_TxReg), активных энхансеров (H3K4me1, H3K27ac) и промоторов (H3K4me3). В меланоцитах дополнительно отмечена ассоциация с фланкирующими активными промоторами (2_TssAFlnk) и открытым хроматином (DNase). В эпидермальных кератиноцитах (NHEK) профиль дополнен меткой активного промотора H3K9ac_Pro и также DNase. Полиморфизм rs1695 расположен в участке, предсказанном для связывания транскрипционных факторов и ко-регуляторов: BAF155, BCL, Pax-4, SIX5, ZID и Znf143. SNPrs1138272 демонстрирует универсальный профиль активного регуляторного хаба во всех основных типах клеток кожи (фибробластах, кератиноцитах, меланоцитах, эпидермальных кератиноцитах), устойчиво ассоциируясь с сигнатурами транскрипции (3_TxFlnk), регуляторных областей (9_TxReg), активных энхансеров (H3K4me1, H3K27ac) и промоторов (H3K4me3). rs1138272 расположен в участке, предсказанном для связывания широкого спектра ключевых транскрипционных факторов и хроматиновых архитекторов: CTCF, E2F, EWSR1, IRF, MAF, MYC, NRSF, POU2F2, RAD21 и SMC3.
Обсуждение. Несмотря на существенный прогресс в генетических исследованиях, многие молекулярные аспекты патогенеза псориаза, в частности роль системы биотрансформации ксенобиотиков и поддержания редокс-гомеостаза, остаются недостаточно изученными. Глутатион S-трансферазы представляют в этом контексте особый интерес, поскольку эта суперсемейство ферментов 2-й фазы биотрансформации ксенобиотиков осуществляет двойную защитную функцию: они не только конъюгируют глутатион с широким спектром эндогенных токсичных продуктов окислительного стресса (таких как 4-гидроксиноненаль), но и играют ключевую роль в детоксикации экзогенных чужеродных химических веществ. Учитывая уже установленное рядом исследований этиологическое значение факторов окружающей среды химической природы [22, 24-28] в развитии и обострении псориаза, изучение генетических вариантов GST приобретает особую актуальность для понимания механизмов предрасположенности и клинической гетерогенности заболевания. В литературе представлены лишь единичные работы, изучавшие ассоциации полиморфизмов генов GST классов theta, mu и pi с риском и проявлениями псориаза. Комплексных исследований, целенаправленно оценивающих вклад широкого спектра генов GST, в особенности тех, что активно экспрессируются в коже, до сих пор не проводилось ни в российской, ни в международной научной практике. Восполняя указанный пробел, настоящее исследование представляет собой первую попытку комплексного анализа роли полиморфных вариантов генов глутатион-S-трансфераз в молекулярных механизмах и клиническом разнообразии псориаза. В фокус исследования были включены гены GSTA4 и GSTP1, кодирующие изоформы GST, наиболее активные в кожных покровах.
Глутатион S-трансфераза A4 катализирует детоксикацию 4-гидроксиноненаля – токсичного продукта перекисного окисления липидов [60]. В настоящем исследовании не выявлено ассоциаций полиморфных вариантов GSTA4 с общим риском псориаза, однако обнаружены их связи с отдельными клиническими фенотипами. Полиморфизмы гена продемонстрировали разнонаправленные эффекты: одни варианты ассоциировались с поражением кожи конечностей, другие – с частотой рецидивов или вовлечением различных суставов, при этом некоторые аллели проявляли протективный эффект в отношении развития псориатического артрита. Функциональные данные указывают на тканеспецифическое влияние изученных SNP на экспрессию GSTA4. Согласно биоинформатическому анализу, ряд полиморфизмов локализован в регуляторных регионах, связывающих транскрипционные факторы (Cdx, AP-1, Hmbox1, Nanog), участвующие в дифференцировке эпителия, клеточной пролиферации и воспалительном ответе [61, 62, 63]. Аллели, ассоциированные со снижением экспрессии GSTA4, могут способствовать локальному накоплению 4-гидроксиноненаля, усиливая окислительный стресс и воспаление в коже или суставах. Напротив, варианты, повышающие экспрессию в фибробластах кожи, проявляют защитный эффект против артрита, вероятно, предотвращая системную диссеминацию воспаления. Наблюдаемый диссоциированный профиль экспрессии (повышение в коже на фоне снижения в крови) может отражать компенсаторный локальный ответ на фоне системной недостаточности антиоксидантной защиты, что создает предпосылки для хронизации воспаления и развития суставного синдрома.
Глутатион S-трансфераза P1 катализирует конъюгацию глутатиона с гидрофобными электрофилами, обеспечивая их детоксикацию [64]. Фермент также участвует в метаболизме простагландинов, биосинтезе провоспалительных липидных медиаторов [65, 66] и негативной регуляции ферроптоза через детоксикацию 4-гидроксиноненаля [67]. В настоящем исследовании выявлены разноуровневые эффекты полиморфизмов GSTP1 на риск и клинические проявления псориаза. Интронный вариант rs1790753, локализованный в сложном регуляторном локусе с тканеспецифичной активностью в клетках кожи, продемонстрировал протективный эффект в отношении поражения кожи туловища и ряда суставов, однако его влияние на общий риск заболевания модулировалось гаплотипическим контекстом. Полиморфизм rs749174, расположенный в высоко консервативном регуляторном модуле, ассоциировался со снижением риска поражения верхних конечностей, ногтей и плечевых суставов, причем его эффект опосредовался факторами среды. Функционально значимые миссенс-варианты rs1695 (Ile105Val) и rs1138272 (Ala114Val), снижающие каталитическую активность фермента, коррелировали с более тяжелыми фенотипами: первый — с площадью поражения кожи нижних конечностей, второй – с развитием артрита голеностопных суставов. Биоинформатический анализ выявил локализацию изученных полиморфизмов в регуляторных регионах, предсказанных для связывания широкого спектра транскрипционных факторов – участников стрессового ответа (NF-kappaB, ATF3), иммунной регуляции, ретиноидной сигнализации (RXRA), контроля клеточного цикла и архитектуры хроматина (CTCF, E2F). Это позволяет рассматривать данные SNP как потенциальные функциональные варианты, способные модулировать экспрессию генов в коже и нарушать скоординированные межклеточные взаимодействия в патогенезе псориаза.
Таким образом, проведенное исследование раскрывает сложную архитектуру генетического контроля псориаза, в которой гены семейства GST выступают не как факторы общей предрасположенности, а как динамические модуляторы клинического разнообразия заболевания. Для генов GSTA4 и GSTP1 показана его роль в определении локализации и характера поражения через регуляцию тканеспецифичного антиоксидантного баланса. Впервые установленная роль полиморфизмов rs749174 и rs1790753 GSTP1 в модуляции предрасположенности к псориазу через диет-зависимые и гаплотипические механизмы дополняет современное понимание патогенеза заболевания. Описанный в литературе повышенный уровень экспрессии GSTP1 в псориатических бляшках [68] может отражать компенсаторную реакцию на генетически обусловленную функциональную недостаточность фермента. Выявленные гено-фенотипические ассоциации полиморфных вариантов генов GST с клиническими параметрами псориаза открывают перспективы для уточнения клинико-патогенетических подтипов заболевания, стратификации пациентов по характеру, распространенности и течению болезни, а также по риску развития внекожных проявлений, в первую очередь псориатического артрита. Полученные результаты создают основу для разработки более персонализированных стратегий терапии и профилактики, учитывающих как генетический профиль пациента, так и модифицируемые средовые факторы.
Заключение. Полученные результаты открывают возможности для уточнения клинико-патогенетических подтипов псориаза на основе генетического профиля пациентов. Перспективным направлением является разработка диагностической панели для прогнозирования индивидуального риска развития псориатического артрита, локализации кожных поражений и характера течения заболевания. Выявленные ассоциации создают основу для стратификации пациентов по вероятному фенотипу болезни, что позволит обосновать более раннее назначение активной терапии пациентам с неблагоприятным генетическим прогнозом. Внедрение результатов генотипирования в клиническую практику будет способствовать переходу к персонализированному ведению пациентов с псориазом, учитывающему не только клиническую картину, но и индивидуальные генетические особенности, определяющие предрасположенность к тем или иным проявлениям заболевания.
Информация о финансировании
Финансирование данной работы не проводилось.














Список литературы