
Pre-treated Enterococcus faecalis with Sub-inhibitory concentrations of Cefepime Modulate Biofilm Formation and Adhesion in vitro
Aннотация
Background: Modifying biofilm formation and adhesion could aid in developing treatments for infectious diseases caused by Enterococcus faecalis, a multidrug-resistant bacteria responsible for urinary tract infections (UTIs). The aim of the study:To investigate the effect of cefepime sub-inhibitory concentrations on the biofilm production and adhesion of E. faecalis. Materials and methods: Seventy-three urine specimens were collected from inpatients with UTIs to isolate E. faecalis. The minimum inhibitory concentrations (MICs) of cefepime were determined to assess the susceptibility of ten E. faecalis isolates. Biofilm formation to polystyrene and adhesion to human oral mucosal epithelial cells (OMECs) in vitro was measured. The effects of sub-MICs of cefepime on biofilm formation, adhesion, and esp gene expression were examined. Results: The present study reported a high incidence of UTI caused by E. faecalis (13.15%). The susceptibility of ten E. faecalis isolates to cefepime and their biofilm formation varied. No significant relationship was observed between the susceptibility of the isolates to cefepime and their biofilm formation (r = -0.28, P = 0.42). Sub-MICs of cefepime (½ MIC, ¼ MIC, 1/8MIC, and 1/16 MIC) reduced biofilm formation and adhesion to human OMECs (P<0.05). Additionally, the pre-treated E. faecalis with above sub-MICs down-regulated esp gene expression (P<0.05). Conclusion: Sub-MICs of cefepime decrease biofilm formation and E. faecalis adhesion to biotic and abiotic surfaces. This effect is achieved through the down-regulation of esp gene expression
К сожалению, текст статьи доступен только на Английском
Introduction.Enterococcus faecalis is a Gram-positive streptococcal bacterium that has raised significant concern in medical microbiology due to its status as a multidrug-resistant pathogen associated with nosocomial infections. It is a part of the normal gut flora; however, it can cause severe endocarditis, urinary tract infection (UTI), and wound infections, particularly in immunocompromised patients [1, 2]. The formation of E. faecalis into biofilm represents a big challenge for curing the infectious diseases caused by this pathogen. It can adhere to biotic and abiotic surfaces in immunocompromised patients and evade host defenses, especially the epithelial cells [3]. Biofilm formation enhances bacterial survival in hostile environments, contributes to persistent infections, and protects the bacteria from antibiotic therapy and host immune responses [4].
Cefepime, a fourth-generation cephalosporin, is usually effective against different Gram-positive and Gram-negative bacteria [5]. It is primarily used to treat uncomplicated urinary tract infections (UTIs). It binds to penicillin-binding proteins (PBPs), disrupting bacterial cell wall synthesis, and leading to cell lysis and death [6]. However, the use of antibiotics at sub-inhibitory concentrations (sub-MIC) has been shown to influence bacterial behavior, including biofilm formation, without necessarily inhibiting growth. Previous investigations have indicated that sub-inhibitory concentrations (sub-MICs) of antibiotics promote bacterial virulence by inducing the expression of biofilm-related genes or facilitating adhesion to different surfaces [7]. Recent studies showed that the sub-MICs of antibiotics reduced the biofilm production and the ability of Pseudomonas aeruginosa to adhere to human epithelial cells in vitro [8, 9, 10]. Biofilm formation and adhesion to biotic surfaces are interesting when studying the pathogenicity of E. faecalis, as these processes can elucidate the mechanisms underlying treatment failures in clinical settings. Biofilm formation enhances pathogenicity and increases bacterial resistance to antibiotic treatment.4 Different genes are involved in the biofilm formation of E. faecalis including esp (the Enterococcal Surface Protein) gene that encodes a surface-associated protein, which is involved in the role of the initial stage of biofilm formation [11].
Understanding how sub-MICs of cefepime affect biofilm formation and adhesion is important for people responsible for managing public health institutions and programs, as it may provide insight into novel treatment strategies and inform clinical practice. E. faecalis possesses unique attributes that allow it to adapt in the presence of antimicrobial agents, such as its capacity for horizontal gene transfer and pseudomonal biofilm architecture, which complicates treatment efforts [12].
Previous studies have primarily focused on the bactericidal effects of cefepime and nitrofurantoin against E. faecalis [13], but limited research has been dedicated to understanding its ramifications on biofilm formation at sub-MIC levels. We propose to investigate how these sub-inhibitory concentrations of cefepime and nitrofurantoin affect the biofilm formation and adhesion to biotic and abiotic surfaces by E. faecalis and we try to suggest the mechanism that regulates these responses.
The aim of the study. To investigate the effect of cefepime sub-inhibitory concentrations on the biofilm production and adhesion of E. faecalis.
Materials and methods
Isolation and Identification
The midstream urine specimens were collected in a sterile container from inpatients (73 samples) suffering from UTIs (Baghdad Teaching Hospital, Baghdad, Iraq). All patients did not receive antibiotic treatment 72 h before the sample collection date and consented to participate in the study (they signed a consent form). The samples were immediately transferred to the laboratory. Briefly, the urine samples were cultured onto MacConkey agar (HiMedia, Mumbai, India), and cysteine lactose electrolyte deficient (CLED) (Himedia) and incubated for 18 h at 37 oC. The bacteria were identified based on appearance on gram stain, growth in 6.5% NaCI, catalase-negative, and growth on bile esculin medium (Hardy Diagnostics). The VITEK 2 DensiCheck instrument (bio- Mérieux, Marcy-l’Étoile, France) was used to confirm the identification of the isolates [14]. The bacterial isolates were subcultured onto the nutrient agar plates for short-term storage [15].
Minimum inhibitory concentration (MIC)
The micro-dilution technique of Al-Mutalib & Zgair (2023) was followed to measure the MICs of cefepime against ten E. faecalis (Ef1, Ef2, Ef3, Ef4, Ef5, Ef7, Ef8, Ef9, Ef10, and Ef18) isolates. A stock concentration of 2 mg/ml was prepared by dissolving 500 µg of cefepime (Sanofi) in sterile Mueller-Hinton broth. Double-fold dilutions (100 μL) were prepared in the microtiter plate (U shape) with sterile MHB (HiMedia). Five microliters of E. faecalis were added to each well. The bacterial suspension was prepared by washing the overnight bacterial growth (cultured at 37 °C in MHB) with sterile phosphate-buffered saline (PBS; 0.1 M, pH 7.2), using centrifugation at 5,000 g for 10 minutes (Beckman Coulter, Brea, USA). The optical density of the bacterial suspension was then adjusted to 0.1 at a wavelength of 600 nm using a double-beam spectrophotometer (model SP-MUV8000T; Bioevopeak, Jinan, China). The plates were shaken gently. Three controls were prepared. MHB and bacterial isolate (1st control), the wells of MHB only (2nd control), and different double dilutions of antibiotics only (3rd control). The plates were incubated at 37 oC for 18 h. The lowest antibiotic concentrations completely inhibit growth considered MIC [8].
Biofilm formation
The previous method was followed to measure the biofilm formation of the ten E. faecalis isolates. Briefly, 200 µl of sterile Tryptic Soy Broth (TSB, HiMedia) were added to flat-bottom polystyrene tissue culture plate wells. The standard inoculum (5 µl) of E. faecalis was added to each well and incubated at 37 oC for 24 h. The bacterial suspension was prepared by washing the overnight growth (grown at 37 oC in TSB) of bacteria with sterile PBS (0.1 M, pH 7.2) using centrifugation at 6,000 g for 12 min (Beckman Coulter, Brea, USA), and the optical density of bacterial suspension was adjusted to 0.1 at a wavelength of 600 nm (Bioevopeak, Jinan, China). The TSB was discarded, and the plates were washed three times with sterile distilled water. The plates were dried (for 30 min at 55 oC). 200 µl of Hucker crystal violet (0.4%) was added to each well. The plates were incubated at room temperature for 15 min and washed 5 times with distilled water. After drying the wells, 200 µ of anhydrous ethanol was added to the wells. The optical density (OD) was measured at a wavelength of 570 nm using a microplate reader (BioTek 800 TS; Winooski, USA). The experiment was repeated in triplicate [10].
Effect of sub-MICs on Biofilm Formation
In this experiment, the effect of different concentrations of cefepime and nitrofurantoin on the biofilm formation of ten clinical isolates of E. faecalis was evaluated. The method of Talib & Ghafil, (2024) was followed to evaluate the level of biofilm formation with modifications. Instead of TSB, the serial dilutions of sub-MICs of cefepime (½ MIC, ¼ MIC, 1/8 MIC, 1/16 MIC, 1/32 MIC, and 1/64 MIC) were prepared in TSB (Himedia) in the wells of a polystyrene microtiter plate (flat shape) were used. The plates were incubated at 37 oC for 18 h and washed thrice with sterile distilled water. The wells were stained with 200 µl of crystal violet (for 15 min) after washing three times with distilled water and drying. The wells were washed five times with distilled water and dried. Two hundred microliters of anhydrous ethanol were added to each well. The absorbency was measured (OD 570 nm) using a microplate reader (BioTek 800 TS, Winooski, USA). The experiment was repeated in triplicate [9, 10].
Effect of antibiotic sub-MICs on adhesion to human OMECs
In this experiment, the E. faecalis (Ef9) that produced the highest level of biofilm was used to study the effect of different concentrations of cefepime (½ MIC, ¼ MIC, 1/8 MIC, 1/16 MIC, 1/32 MIC, and 1/64 MIC) on E. faecalis adhesion to human OMECs. The previous methods were followed to prepare human oral mucosal epithelial cells (OMECs) in vitro and to check the effect of different concentrations of cefepime on the adhesion of E. faecalis (EF9) to human OMECs.8-10,16 The bacterial colonies were grown onto TSB (Himedia). The standard inoculum of bacteria (1×107 c.f.u./ml) was prepared in TSB and treated with the different concentrations of cefepime (½ MIC, ¼ MIC, 1/8 MIC, 1/16 MIC, 1/32 MIC, and 1/64 MIC) and incubated for 18 h at 37°C. The bacteria suspensions were washed 3 times with PBS (0.1 M, pH, 7.2) by centrifugation at 10,000 g for 5 min. The bacterial cells were re-suspended in TSB to check the ability of pre-treated bacteria with cefepime to adhere to human OMECs. In tissue culture tubes (BIOFILR, China) 1×105 OMECs were suspended in Dulbecco’s modified Eagle’s medium (D-MEM) containing 10% fetal calf serum, 10 mM L-glutamine and 100 μl of E. faecalis (5×107 c.f.u./ml) were mixed. The tubes were incubated at 37 oC for 2 h. Then the OMECs were washed thrice with PBS (0.1 M, pH, 7.2) using centrifugation at 1,000 g for 10 min (Beckman Coulter, Brea, USA). The human OMECs were lysed with PBS-0.5% Triton×100 (Sigma-Aldrich). The suspension was diluted tenfold and plated on nutrient agar to count the viable bacteria. The untreated bacteria with cefepime were considered a control group [9, 16].
Gene detection and Real-time PCR
The method reported by Caixeta Magalhães Tibúrcio et al. (2022) was followed to detect the esp gene in the Ef9 (E. faecalis that produced the highest level of biofilm) [17]. The standard method of Van Wamel et al. (2007) was followed to evaluate the esp gene expression in the Ef9 isolate [11]. The RNA was isolated from bacterial suspension (0.5x108 c.f.u./ml) according to the procedures of Cheung et al. (1994) [18]. The RNA purification and cDNA generation were done as described by Nallapareddy et al. (2006) [19]. The Real-time PCR was performed using TaqMan Universal PCR master mix (Applied Biosystems). The chosen PCR conditions were described previously.11 The ΔΔCT method of Livak et al. (2001) was used to calculate the difference in esp mRNA present in bacteria grown at 37°C and at 21°C using 23S rRNA as an internal control [20]. Analyses were performed in triplicate.
Statistical analyses
The statistical analysis was conducted and the graphs were generated utilizing Origin v. 8 software (OriginLab, Northampton, USA). The data were presented as means ± standard error (M ± SE). The disparities were assessed utilizing a Student’s t-test and one-way analysis of variance (ANOVA). The relationship was evaluated with Pearson’s correlation coefficient (r). A P< 0.05 was deemed statistically significant [21, 22].
Results
Bacterial Isolates
Ten isolates of E. faecalis were isolated from 73 urine samples collected aseptically from patients suffering from UTIs. The bacterial species were identified using microscopic and biochemical tests. The isolates were further confirmed as E. faecalis using VITEK 2 technology. The present study revealed a high incidence of urinary tract infections with E. faecalis (13.15%).
Antibiotic Susceptibility and Biofilm Formation
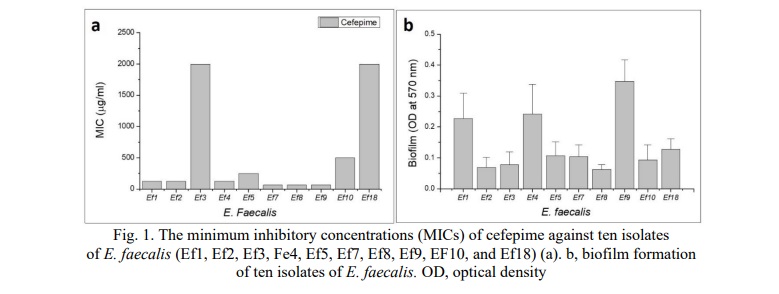
The MICs were measured for ten isolates of E. faecalis against cefepime. The results showed that the highest MIC of cefepime was against Ef3 and Ef18 (2000 µg/ml) followed by Ef10, while the lowest MIC of cefepime was against Ef7, Ef8, and Ef 9 (62.5 µg/ml) (Fig. 1a). Fig. 1b shows the levels of biofilm of ten isolates of E. faecalis. The E. faecalis (Ef9) produced the highest level of biofilm (0.34±0.06) followed by Ef4 (0.24±0.096). The lowest biofilm was produced by Ef8 (0.062±0.015). The study showed that the isolates produced the biofilm on a polystyrene microtiter plate. That is why the isolate Ef9 will be used in further experiments.
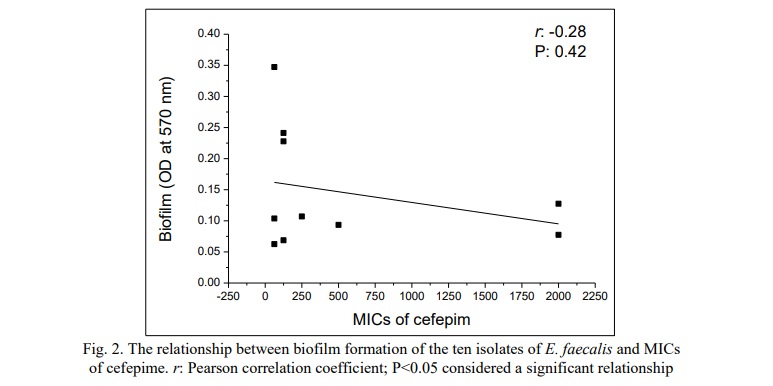
The relationship between the MICs of cefepime against ten isolates of E. faecalis that were isolated from urine specimens collected from patients with UTI and biofilm formation of the isolates was investigated. The study showed there is no statistical correlation between the MIC of cefepime (r:-0.28, P>0.05) against ten isolates and the biofilm formation of the same isolates (Fig. 2).
Influence of Sub-MICs of Cefepime on Biofilm Formation
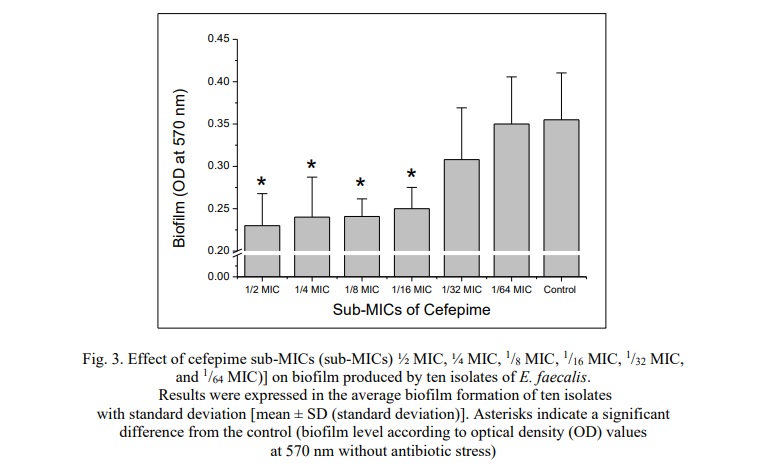
Effects of the sub-inhibitory concentrations of cefepime on the biofilm formation of the isolates of E. faecalis (10 isolates) on polystyrene were evaluated. The results were expressed as the mean ± standard deviation of the biofilm formation (OD570nm) of the ten isolates of E. faecalis. The current study demonstrated that sub-MICs of cefepime (½ MIC, ¼ MIC, 1/8 MIC, and 1/16 MIC) significantly reduced biofilm production (P<0.05) compared to the control (biofilm formation of E. faecalis without exposure to cefepime). The treatment of the isolates with 1/32 and 1/64 of cefepime did not significantly affect (P>0.05) the biofilm development of the ten isolates of E. faecalis on polystyrene (Fig. 3).
Effect of Cefepime Sub-MICs on adhesion to human OMECs
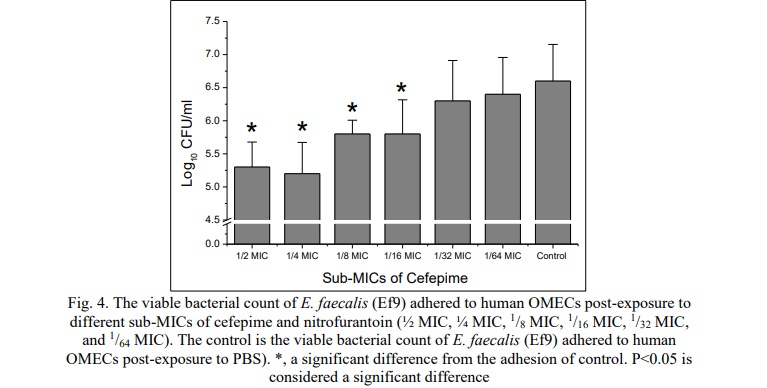
In the current study, the human OMECs were used as a model of the biotic surface to evaluate the effect of different sub-inhibitory concentrations of cefepime (½ MIC, ¼ MIC, 1/8 MIC, 1/16 MIC, 1/32 MIC, and 1/64 MIC) on the ability of Ef9 (the E. faecalis that produced the highest level of the biofilm formation) to adhere to biotic surfaces. The viable bacterial count was used to estimate the number of live bacteria that adhered to the human OMECs (Fig. 4). The results were compared with the viable bacterial count of Ef9 that adhered to human OMECs without antibiotic stress. The study showed a significant decline in bacteria adhesion when the bacteria were pretreated with ½ MIC, ¼ MIC, 1/8 MIC, and 1/16 MIC of cefepime. The present study showed that the effect of sub-MICs of cefepime on the adhesion of E. faecalis to epithelial cells is variable and dependent on the sub-MICs.
Effect of Sub-MICs of Cefepime on esp Gene Expression
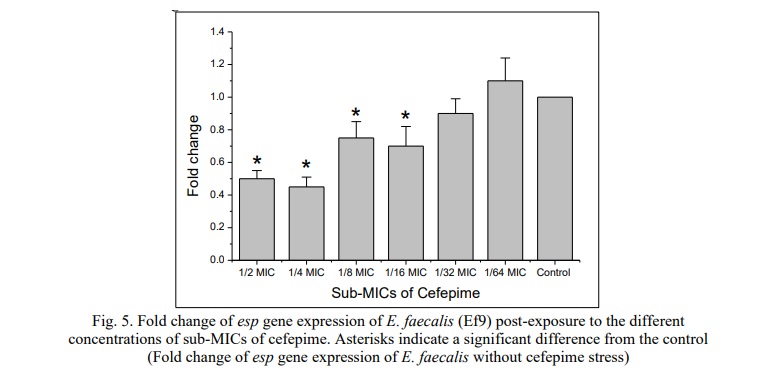
The study demonstrated the effect of sub-MICs of cefepime on esp gene expression. This gene is involved in the initial stage of biofilm production. The experiment was conducted on E. faecalis (Ef9), which produced the highest level of biofilm on the polystyrene microtiter plates. Fig. 5 shows that sub-MICs of cefepime (½ MIC, ¼ MIC, 1/8 MIC, 1/16 MIC) significantly down-regulated the esp gene expression (P<0.05). However, no significant change in esp gene expression was observed when the Ef9 was pre-exposed to 1/32, and 1/64 MICs of cefepime (P>0.05).
Discussion. The formation of biofilm by pathogenic bacteria is a significant factor contributing to their persistence and virulence, particularly in nosocomial infections that exhibit resistance of bacteria to a wide spectrum of antibiotics. Earlier studies have shown that sub-inhibitory concentrations of antibiotics variably affect bacterial biofilm formation [8, 9, 10, 16, 17]. Investigating the role of such concentrations of cefepime in modulating E. faecalis biofilms may provide appreciated insights into developing more effective treatment strategies and the effect of low concentrations of antibiotics in curing E. faecalis infectious diseases including UTIs [23].
In the present study, ten E. faecalis isolates were isolated from the urine of patients with UTIs. The incidence of infection with these bacterial isolates among UTI patients was relatively high. The susceptibility of the isolates to cefepime in terms of the MICs of cefepime was assessed, revealing variable responses among the isolates to the antibiotic. Similarly, the biofilm-forming ability of the isolates also exhibited variability. No significant relationship was observed between the biofilm formation of the isolates and their susceptibility to cefepime. The study further demonstrated that exposure to sub-inhibitory concentrations of cefepime reduced the biofilm formation on the abiotic surface (polystyrene) and adhesion to biotic surfaces (human OMECs) of the isolates, particularly at higher sub-MIC levels. Additionally, higher sub-MICs of both antibiotics were found to down-regulate the expression of the esp gene.
No previous studies investigated the effect of sub-inhibitory concentrations of cefepime and nitrofurantoin on the biofilm formation of E. faecalis. Caixeta Magalhães Tibúrcio et al. (2022) reported that the sub-MICs of ampicillin, penicillin, and gentamicin interfere with the expression of biofilm formation genes of E. faecalis and they suggested this topic required further study to clarify the role of sub-MIC of the antibiotics on the biofilm formation of E. faecalis [17]. Bernardi et al., (2021) investigated the effect of sub-inhibitory concentrations of Penicillin G, Amoxicillin, Doxycycline, Fosfomycin, Tetracycline, and Vancomycin on the biofilm formation abilities of E. faecalis, they found that the sub-MICs of the studied antibiotics enhanced the biofilm formation in vitro [24]. In contrast, previous studies reported that the sub-MICs ofloxacin, rifampicin, and ceftriaxone reduce the biofilm formation of P. aeruginosain vitro [8, 9, 10].
The reduction in biofilm formation and adhesion of bacteria observed in this study may be attributed to the interference of antibiotics with key mechanisms involved in biofilm formation, such as quorum sensing (QS), gelatinase (GelE), cytolysin, enterococcal surface protein (Esp), and the role of pili [17, 25, 26, 27]. The findings highlight that sub-MIC levels of cefepime interfere with biofilm formation, suggesting their potential as adjunct therapies to combat biofilm-associated infections and improve treatment outcomes in UTI patients. Further research is necessary to clarify the mechanisms of sub-MIC levels to reduce biofilm formation and to assess their impact on bacterial susceptibility to antibiotics. While our study provides valuable insights, reliance solely on in vitro results is insufficient; in vivo studies are essential to validate and extend these findings [28].
This study also opens avenues for future investigations into the effects of low concentrations of cefepime on bacterial virulence, particularly their biofilm-forming ability, under both in vitro and in vivo conditions. Additionally, exploring the influence of these antibiotics at different stages of biofilm formation could provide a deeper understanding of their therapeutic potential.
Conclusion. The high incidence of UTIs caused by E. faecalis is well-documented. The susceptibility of clinical isolates of E. faecalis to cefepime is variable. All the E. faecalis isolates studied were capable of forming biofilms on polystyrene microtiter plates. However, their ability to form biofilms was not correlated with their susceptibility to cefepime. The sub-MICs of cefepime reduced biofilm formation and E. faecalis adhesion to human (OMECs) biotic surfaces. Antibiotics interfere with biofilm formation through various mechanisms. In this study, sub-inhibitory concentrations of cefepime down-regulated the expression of the esp gene, providing insight into the mechanism by which cefepime inhibits biofilm formation.
Financial support
No financial support has been provided for this work.
Благодарности
The authors thank all staff members of the Department of Biology, College of Science, University of Baghdad, for all support during the experimental work














Список литературы