
Immunomodulatory effect of saponins from Agave brittoniana subsp. brachypus (Trel.) in a model of fetal alcohol toxicity
Aннотация
Background: Prenatal exposure to ethanol can result in immunosuppression in the foetus due to alterations in tissue architecture and cellular composition, as well as reduced immune system function. This highlights the urgent need to identify new immunomodulatory agents. A variety of natural products and their metabolites are a significant source of molecules with such properties. Agave brittoniana subsp. brachypus (Trel.) A. Álvarez is a species known to produce steroidal saponins, which have demonstrated immunomodulatory activity. The aim of the study: To examine the immunomodulatory effects of saponins derived from the A. brittoniana species. Materials and methods: A fetal alcohol toxicity model was developed. Progenitor rats were orally administered 40 % alcohol (5 g/kg/day on gestational days 3-21), followed by treatment with three doses of saponins-enriched extracts (25, 50 and 100 mg/kg bw) and folic acid, which served as a control. The cellular and humoral response of the immune system was evaluated in neonates at 15, 21 and 31 days after birth. Results: In neonates with fetal alcohol toxicity, an increase in leukocytes at the expense of lymphocytes was observed in the group with the highest dose of saponins. The humoral response of these neonates was characterised by increased IgG at all three doses, with a restoration of C3 values only observed in the 100 mg/kg A. brittoniana saponin group. Conclusion: Steroidal saponins from A. brittoniana species have high immunomodulatory potential in immunocompromised animals
К сожалению, текст статьи доступен только на Английском
Introduction. Immunomodulators are substances that modify the ability of the immune system to exert one or more of its functions, including antibody production, antigen recognition, and the secretion of inflammatory mediators. Their activity indicates a biological or pharmacological effect on humoral or cellular factors involved in the immune response. The stimulation or suppression of these factors may contribute to the maintenance of good health [1].
It is estimated that millions of individuals worldwide are afflicted with diseases that compromise the immune system, including cancer [2]. The use of immunomodulatory drugs provides an additional therapeutic tool to conventional therapy in individuals with impaired immune function. Furthermore, they are employed in the treatment of inflammatory diseases, allergic conditions, diabetes mellitus, acquired immune deficiency syndrome (AIDS), as well as viral infections that affect the immune system, including the COVID-19 pandemic [1, 3].
Alcohol consumption represents the third leading risk factor for disease and disability globally. In the Americas, it is the primary contributor to these health issues. The mean quantity of alcohol consumed per capita in Cuba is less than the regional mean for the Americas, as indicated in reports from 2018 [4].Conversely, there has been a disconcerting global surge in alcohol intoxication among young women and women of reproductive age, a phenomenon that has also been observed in our country [5].
Although studies of alcohol-induced toxicity have traditionally focused on the neurological, hepatic and gastric damage that it causes, the negative effect on the immune system is also known[6].Immune system disorders are a common occurrence in the paediatric age group, with some evidence suggesting a potential link to prenatal alcohol consumption [7].
Fetal alcohol syndrome (FAS) is a condition characterised by a range of physical, cognitive and developmental disorders resulting from prenatal alcohol exposure. Immunosuppression results from the direct and indirect exposure of the developing immune system to the toxic alcohol metabolites acetaldehyde and acetate. This exposure leads to persistent alterations in tissue architecture, cellular composition, and the immune system's functional capacity [8].
Chronic alcohol consumption has been associated with increased levels of proinflammatory cytokines [9]. This finding suggests that chronic alcohol consumption during pregnancy may expose the foetus to an increased oxidative environment and dysregulation of inflammatory responses. This increase may compromise the integrity of fetal innate and acquired immunity [10].
The diminished innate response in the FAS is reflected in impaired macrophage and neutrophil function, partly due to mitochondrial damage caused by intense oxidative stress and glutathione depletion [11]. It has been demonstrated that immune dysfunction can result in a decline in lymphocyte levels. This stage is consequent to interference with signalling pathways associated with lymphocyte differentiation and activation [12]. Consequently, lymphopenia and an imbalance of critical subpopulations, which impact on humoral immunity with decreased immunoglobulins, as well as reduced cellular response due to decreased leukocyte subpopulations, are pathognomonic for immune dysfunction in the context of FAS [12, 13].
It has been demonstrated through experimental studies that ethanol and its metabolites can induce alterations in the processes involved in the selection of T-lymphocyte within the thymus. Ethanol-induced immune dysfunction has also been accompanied by a reduction in the number of CD4+ and CD8+ clones present in the peripheral blood [14, 15]. Consequently, numerous biomedical researchers are engaged in the pursuit of novel, less toxic, plant-derived compounds with the potential to stimulate the immune response in immunocompromised patients [16, 17].
The search for natural products and their components represents a significant source of discovery in the field of immunomodulatory drugs and their novel mechanisms of action. In this context, a considerable number of studies have been published demonstrating the immunomodulatory activity of plants such as Boerhaavia diffusa (Nyctaginaceae), Curcuma longa (Zingiberaceae), Asparagus racemosus (Asparagaceae), Panax ginsengs (Araliaceae), Arnica montena (Compositae), Echinacea purpurea (Asteraceae)and Calendula officinalis (Asteraceae) [18].
The species Agave brittoniana Trel. subspecies brachypus (fam. Asparagaceae), described by Trelease in 1913 and updated by Álvarez de Zayas [19] has been the subject of phytochemical studies. Agave brittoniana T contains significant quantities of glycosylated steroidal saponins [20, 21]. The presence of these metabolites has been associated with the plant's anti-inflammatory potential in acute and chronic models [22, 23] as well as in ulcerative colitis model in rats [24]. However, these compounds have not been studied in relation to their effect on the immune system. Nevertheless, in other species where they are present, this property has been demonstrated [25].
Plant extracts with high saponin content have been demonstrated to regulate both innate and acquired immune responses [26]. Saponin-based adjuvants stimulate cell-mediated immunity and enhance antibody production. They also have the advantage that only low doses are required for adjuvant activity. Research has demonstrated that these metabolites, when utilised alone or in conjunction with aluminium salts, liposomes or lipid/protein complexes, function as immunostimulants and adjuvants within vaccine formulations [27].
Saponins have been demonstrated to regulate a variety of immune cells through the secretion of related cytokines and specific antigens. Activation of macrophages and natural killer cells is necessary for the neutralisation of harmful agents and subsequent release of pro-inflammatory cytokines that contribute to the amplification of the immune response [26]. This phenomenon is promoted by saponins, such as those extracted from Panax notoginseng species [28] or platycodin D extracted from the stem bark of Albizia julibrissin [29].
A number of studies have indicated that saponins possess anti-inflammatory properties, which may be attributed to their capacity to suppress the secretion of cytokines and acute phase reactants, which are involved in the inflammatory response [28]. As has been demonstrated in multiple studies, an inflammatory environment occurs during the immune response due to the reaction of components of the immune system to recognise and inactivate antigenic molecules[3].It is therefore logical to conclude that molecules with an effect on these mediators could modulate the immunogenic response.
The present research explores the immunomodulatory potential of the species' saponins for the first time. Consequently, the research seeks to assess the impact of these molecules on the leukocyte population and antibodies, as the most visible expression of innate and acquired immunity.
The aim of the study. To examine the immunomodulatory effect of saponins-enriched extracts (SEE) from the species A. brittoniana in an experimental model of fetal alcohol toxicity (FAT), which results in the development of immunodeficiency in neonates.
Materials and Methods. The present study evaluated the immunomodulatory effects of three dose levels of A. brittoniana saponins, isolated through butanolic fractionation, in neonatal rats subjected to FAT. The leaves of A. brittoniana were collected from the Cubanacán protected area, located on the periphery of Santa Clara city. The botanical identification of the species was conducted at the Research Center of the Botanical Garden, Central University “Marta Abreu” of Las Villas, where the specimen was matched with the serial number HPVC 5445.
Obtaining the saponins-enriched extracts (SEE)
Leaves exhibiting optimal phytosanitary conditions (absence of mechanical or chemical damage) were selected for subsequent processing. Following the methodology established in previous work [22], they were first dried and pulverised. The finely cut plant material was dried for 7 days at 40 °C in an oven. The dried material was then milled in a cutting mill (IKA-MF 10B, Germany) at 3000 rpm. The resulting product was then sieved through a sieve with an internal diameter of 3 mm.
In order to obtain the hydroalcoholic extract, 300 g of the dried leaf powder were extracted three times with 1 L of 70 % ethanol by maceration at room temperature in the dark conditions. The solvent was removed by using an IKA® RV 10 basic rotoevaporator at 40°C and steam pressure. The syrupy extract (10 g) was resuspended in a single portion of distilled water, placed in a separatory funnel and 36 ml of water-saturated n-butanol was added. The mixture was left for 24 hours and then the organic phase was concentrated under steam pressure at the conditions previously mentioned [22].
Saponin predominance was corroborated by phytochemical screening in accordance with the methodology proposed by Miranda and Cuellar [30]. In addition, the SEE were evaluated on thin-layer chromatography (TLC) using methanol:chloroform (3:7) as the mobile phase, and then 50 % H2SO4 was used as a developer. The products of the saponin-enriched extracts were compared with the diosgenin standard and purified yucagenin (from the UCLV Organic Chemistry Laboratory), in line with prior studies that identified these steroidal compounds as the principal aglycons present in Agabrittonosides (steroidal saponins of Agave brittoniana species) [20, 21].
Animals
Wistar rats were obtained from National Center for Laboratory Animal Production (CENPALAB Spanish acronym) and maintained under controlled environmental conditions (12-hour light/dark cycle, 20±2°C, 50 - 70 % relative humidity), with conventional feeding and water ad libitum. The study involved thirty-six Wistar rats (with weights ranging between 180 and 200 g) and sixteen males weighing between 200 and 270 g, which served as the progenitor population. The first day of gestation was determined by the presence of spermatozoa in the vaginal smear in the early morning hours.
Experimental Model of Fetal Alcohol Toxicity
The ethanol administration scheme was designed to take into account the previously reported toxic effects on prenatal alcohol exposure [31, 32].The model reported by Wang et al. was modified to achieve a toxic effect of ethanol on maternal rats without causing mortality or serious teratogenic effects [33].
A total of six experimental groups were established, each consisting of six dam rats (n=6). The groups were designated as follows: I: healthy group, lacking the induction of FAT, without any further intervention II: negative control, FAT induction without treatment, III: FAT induction with folic acid administration at a dose of 5 mg/kg bw. This was followed by treatment with SEE at 25 mg/kg bw IV, 50 mg/kg bw in group V and 100 mg bw in group VI.
Induction of the model and treatment was initiated on the third day post-mating, continuing until parturition in a single daily dosage. Ethanol 40 % was administered orally to all pregnant rats in groups II to VI at a dose of 5 mL/kg bw. Two hours later, the respective oral treatments were initiated.
The sample for immune response studies consisted of 72 male neonates, 12 from each group. Blood samples were collected at 15, 21 and 31 days of postnatal life for the purpose of cellular and humoral immunological studies. The pups were maintained with their mothers until weaning, which occurred 21 days after birth.
Subsequently, the subjects received unrestricted access to water and diet for their specific species until 31 days post-natal life. Upon reaching this age, the subjects were euthanised via an overdose of an inhalational anaesthetic, which was followed by a cervical dislocation.
Assessing the development of immune system dysfunction.
In order to ascertain the effect on the immune system of neonates exposed to ethanol prenatally, the cellular response was quantified by specific techniques for each determination, using heparin as an anticoagulant [34]. To assess potential dysfunction in the humoral response, serum samples were centrifuged at 3500 rpm for 10 minutes to quantify immunoglobulins (IgA, IgM and IgG) and complement fractions (C3 and C4). The quantification techniques were performed using an immunoturbidimetric method [35] provided by Roche, on a Roche/Hitachi Autoanalyser – cobas c 311. In all biochemical determinations, the PreciControl ClinChem Multi 1 and 2 controls were employed, along with the Calibrator f.a.s. Proteins (C.f.a.s.) calibrator.
Statistical processing of the data
The data were subjected to statistical processing using the statistical software program SPSS for Windows, version 22.0. The mean and standard deviation were calculated for each variable under investigation. The Kruskal-Wallis test was employed to ascertain whether there were any statistically significant differences between the groups in each of the experiments. The Mann-Whitney test was employed to ascertain whether there were statistically significant differences between the groups. The variation in the parameters before and after treatment was evaluated using the Wilcoxon test. The following significance levels were considered: p<0.05 (significant) and p<0.01 (highly significant).
Results. In the study evaluating neonates prenatally exposed to alcohol toxicity, blood samples were collected at 15, 21, and 31 days postpartum, corresponding to the lactation, weaning, and postnatal developmental stages. Analysis revealed no significant differences in cellular or humoral immune responses between groups at 15 days, suggesting that prenatal alcohol exposure does not induce immediate immune system impairment during this early postnatal phase.
The results of the cellular immune response during the weaning stage and 31 days postnatal life are presented in the following Table 1.
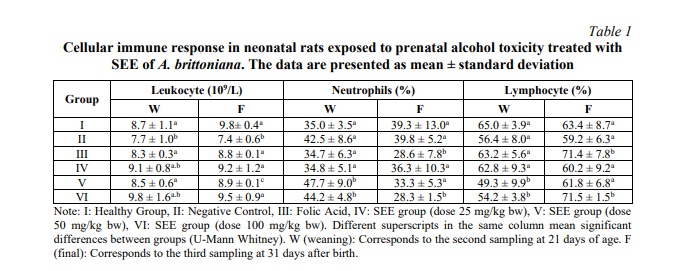
The experimental data show significant changes in the parameters under investigation at 21 and 31 days post-weaning and post-natal, respectively. Despite eosinophils and monocytes being part of the differential leukocyte count, the table above does not include their values. This is because the figures found were not statistically significant. The low cell count for these lineages remained within the normative range for the species: monocytes ranged from 0 to 5, and eosinophils from 1 to 5.
Comparative analysis between the healthy and the negative control group revealed a reduction in the number of leukocytes at 21 days (p=0.001). This finding provides evidence that alcohol consumption is associated with an immunosuppressive response.
Prenatally ethanol-exposed group receiving folic acid supplementation combined with low-dose of SEE exhibited no significant alteration in the cellular response of neonates. In contrast, medium and high-dose SEE groups demonstrated increased neutrophil values (p=0.024 and p=0.009, respectively) and decreased lymphocyte values (p=0.024 and p=0.026, respectively).
While the cellular response in plant saponin-treated groups (21 days post-exposure) failed to demonstrate significant activation relative to the healthy control group, all treated groups exhibited a consistent preventive trend against leukocyte reduction, which was recorded in the negative control group Furthermore, in the case of the 25 and 100 mg/kg doses, the leukocyte values were statistically higher than in the untreated group (p=0.013 and p=0.048, respectively).
After 31 days, it was notable that all groups treated with different doses of SEE from A. brittoniana and folic acid showed a significant increase in leukocytes compared to the negative control group (p = 0.05). A marked increase in lymphocyte values was observed in groups treated with folic acid (p = 0.008 in both cases) and the higher dose of SEE (p = 0.043 and p = 0.002, respectively), compared to the healthy group and negative control group.
Table 2 illustrates the behaviour of the immunoglobulins in the experimental model. The values of immunoglobulin IgA are not displayed in the table as they fall below the detection limits of the test.
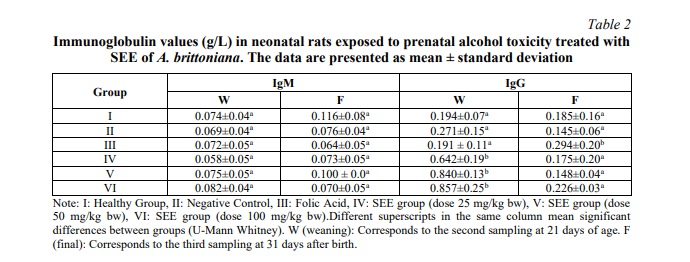
A comparison of the healthy group at 21 days with the remaining groups revealed a significant increase in IgG values in all groups treated with SEE of A. brittoniana at doses of 25 mg/kg (p=0.008), 50 mg/kg (p=0.036) and 100 mg/kg (p=0.036). Furthermore, higher levels of this immunoglobulin were observed in the SEE-treated groups when compared to the negative control (p < 0.05).
Table 3 illustrates the behaviour of serum complement in neonatal rats exposed to prenatal alcohol toxicity.
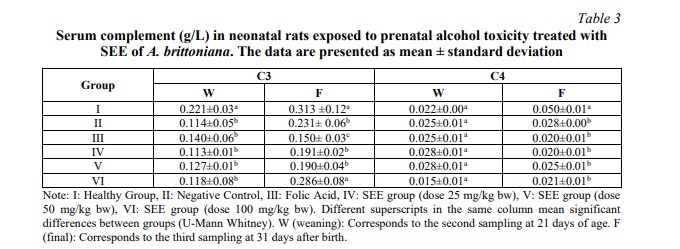
A significant decrease in C3 values was observed in the negative control group after 21 days (p=0.01), as well as in groups treated with SEE of A. brittoniana (p<0.05) and folic acid (p=0.009). No significant differences were observed in complement C4 at 21 days when the experimental groups were compared.
At the conclusion of the study (31 days postpartum), a reduction in C3 and C4 levels was observed in almost all of the experimental groups compared to the healthy group. Doses of 100 mg/kg of SEE produced values that were comparable to those of the healthy group (p > 0.05), with levels of this fraction recovering by the end of the study.
Discussion. Prenatal exposure to alcohol can result in a broad range of physical and behavioural abnormalities. These are categorised into a particular pattern of birth defects known as fetal alcohol syndrome [35].
Pregnancy is a period of immunosuppression in women due to the necessity for the mother's immune system to tolerate the foetus. In this context, it has been demonstrated that the consumption of alcohol, specifically ethanol, and its metabolite, acetaldehyde, has the capacity to interfere with fetal development. The mechanisms by which these substances exert their detrimental effects include the disruption of cell differentiation and growth, the alteration of DNA and protein synthesis, and the inhibition of cell migration. Furthermore, both modify the intermediary metabolism of carbohydrates, proteins and fats, decrease the transfer of amino acids, glucose, folic acid, zinc and other nutrients across the placental barrier, and indirectly affect fetal growth due to intrauterine nutrient deprivation [36, 37].
The outcomes demonstrated in the present study are in accordance with observations recorded in human clinical practice, occurring independent of any infectious agent or allergenic triggers. The immune system may be affected by alterations in neutrophil polymorphonuclear cell mobility and adhesion, as well as in the late hypersensitivity response or type IV. Nevertheless, these values remain within the physiological range reported for the species [38].
Cellular immune response results demonstrate various agave dosage groups potentially counteract alcohol-induced leukocyte reduction [36]. Furthermore, the increase in lymphocytes observed in both the group administrated the highest dose of SEE and folic acid suggests that cell proliferation may be activated, thereby enhancing the immunogenicity of the antigenic proteins [37]. This stage, in turn, makes the antigen-presenting action of the macrophages more effective
Among the principal secondary metabolites isolated from A. brittoniana T. subsp. brachypus are saponins, which have demonstrated immunomodulatory activity in other species containing this metabolite [25, 27]. In studies with rats immunised with a suspension of ram red blood cells, where hydroalcoholic extracts of Phenax rugosus plant containing saponins in its phytochemical composition were administered, there were statistically significant increases in total leukocyte and lymphocyte counts (p<0.05). Other plants that also contain saponins among their secondary metabolites, such as Solanum dolichosepalum also demonstrated increases in leukocyte, lymphocyte and monocyte counts [39]. Sheng et al. [40] observed that the immune system of rats was stimulated by the increase in total leukocyte count observed after 8 weeks of supplementation with cat's claw (Uncaria tomentosa) containing saponins. In a shorter time frame (21 days), the crude saponins from A. brittoniana T. (25 and 100 mg/kg) were observed to increase plasma leukocyte levels.
Guerra and collaborators [20, 21] had proved that the butanolic fraction of the hydroalcoholic extract of A. brittoniana, obtained following the same scheme described in this research, contain steroidal saponins as its major metabolites together with reducing sugars. Other metabolites have been detected in butanolic fraction but at lower concentrations, in different species of agave genus [41, 42].
The negative control group, which was exposed to alcohol toxicity from prenatal age and without treatment, demonstrated a decrease in complement C3 and C4. Maternal alcohol consumption results in a reduction in the blood concentration of various micronutrients, including potassium, magnesium, zinc, phosphorus, calcium, and others. Additionally, the generation of free radicals caused by alcohol intake can lead to structural and functional damage to cells. These alterations, which occur through placental communication, have consequences for the formation and maturation of fetal structures during the gestational period and also during breastfeeding. The intracellular calcium and magnesium levels depressed by the effects of alcohol would modify numerous cell signalling cascades, including the complement system [7, 36].
In this research, we utilised neonatal rats corresponding to human developmental stages of 3, 5, and 7 months of age [43]. Consequently, the figures may be undetectable due to the young age of the biological models. It is critical to note that the synthesis of immunoglobulin A (IgA) initiates late in life, with serum concentrations increasing progressively until achieving normative titters at 4-5 years [44].
This immunoglobulin may partially explain the reduced complement fraction levels observed, as these factors are essential for activating complement cascade intermediates. Conversely, it has been demonstrated that ethanol and acetaldehyde, the primary metabolites of alcohol, alter the hepatocyte cell membrane, thereby activating the complement cascade – particularly the C5a fraction, which is a potent chemotactic factor for neutrophils. The accumulation and activation of neutrophils generates the production of free radicals, which attack molecules in their environment [45]. Consequently, complement levels are decreased due to their consumption in an environment that promotes cellular inflammation. The net result is a marked decrease in complement levels and an increase in the cytotoxic effect of alcohol on the body's own cells, which activates a vicious cycle of increased oxidative stress [37].
The reduced levels of complement factors C3 and C4 were maintained after 21 days in groups treated with different doses of the plant extract, as well as in groups treated with folic acid. The group that received the highest dose of saponins demonstrated the most promising results, with C3 values approaching those of the healthy control group after 31 days. The remaining subjects exhibited continued decreases in C3 and C4 values. This result may indicate that this dose is more effective in neutralising the toxic effects of alcohol, particularly in relation to the inflammatory environment. This behaviour could result in the activation of complement as a consequence of cell damage and a decrease in the synthesis of these protein intermediates, which are affected by direct damage to the hepatocyte where they are synthesised [14, 15].
In the context of the acquired immune response, elevated IgG levels are a predominant phenomenon. This result is consistent with the activation of the complement system, which facilitates phagocytosis [44].Therefore, saponins may have an immunostimulatory effect by increasing IgG production [27].
Saponins have the capacity to stimulate plasma cell receptors, which in turn stimulate the Th1 response. This process results in the production of IFNγ (interferon) and TNFα (tumour necrosis factor), which in turn induces the production of the immunoglobulin IgG. Saponins, due to their lipid nature, may be able to directly stimulate B cells, thus behaving as T antigen-independent [27].
At the conclusion of the study, the increase in IgG observed in the neonates following the administration of saponins to the progenitor mothers was reversed, with the parameter returning to values comparable to those observed in the healthy group. This phenomenon could be explained by the clearance of saponins from the neonates' bodies. It is understood that after 21 days of age, they are weaned and only the saponins passed during the lactation stage remain circulating.
The literature contains publications that address the mechanisms of action that explain the immunomodulatory activity of saponins and justify their potential use in both therapeutics and vaccine production [17, 41].
The complement system is involved in the modulation of the specific humoral immune response. The C3 component of the complement system is the most significant and prevalent protein. The concentration is reduced in a number of cases, including but not limited to inflammatory and infectious diseases, alcoholic patients and environments that inhibit the synthesis of cascade intermediates [38].
The impact of the SEE of A. brittoniana on the immune system of animals with impaired immunity due to alcohol toxicity allows us to observe an increase in the leukocyte response at the expense of lymphocytes at the conclusion of the study. With regard to the humoral response, a dose-dependent increase in IgG can be observed prior to weaning. However, in the group receiving the highest dose, the levels of C3 and C4, which are reduced in the alcohol toxicity model, only recovered to a level similar to that of C3 in the healthy group. C4 remained at lower values. These results suggest that the natural product being investigated could reverse some of the adverse effects associated with alcoholic immunosuppression. Elevating lymphocyte levels and antibody production improves immune system responsiveness to levels comparable with those achieved through folic acid treatment [44].
The low activation of the immune system after 31 days post-exposure may suggest that a different dosing schedule, which takes into account the postnatal age of the subjects, should be employed to assess the consistency of the immunomodulatory effect of the species' saponins in the FAT model.
The preliminary evaluation of immunomodulatory compounds is typically oriented towards non-specific mechanisms that activate both innate and acquired immunity. Consequently, in vivo models are utilised to evaluate the protective capacity of these compounds against immune system injury. Further studies should investigate the type of lymphocyte fraction involved, as well as the specific immunomodulatory mechanism taking into account the type of cytokines involved.
Conclusions. The immunomodulatory capacity of saponin-rich fractions of A. brittoniana has been demonstrated by increasing the cellular and humoral immune response of animals immunocompromised by fetal alcohol toxicity.
Financial support
This research was funded by the project 'Evaluation of the immunomodulatory and antineoplastic potential of Agave brittoniana species' (22-002P/F), part of the National Programme of Natural and Traditional Medicine of the Cuban Ministry of Health.
Conflict of interests
The authors have no conflict of interest to declare.
Благодарности
The authors would like to thank Yuliet González Madariaga, English Language Specialist at the Department of English Language, Faculty of Humanities, Central University “Marta Abreu” of Las Villas, for her expert linguistic review of this work. Her meticulous analysis of the English manuscript and expert suggestions significantly enhanced the linguistic quality and clarity of our scientific presentation. The professional language editing service was essential in ensuring the precision required for effective international academic communication














Список литературы
Список использованной литературы появится позже.