
Anti-Proliferative Activity of [Cu(phen)(D-threo)]NO₃ + F⁻ Against Colorectal and Ovarian Cancer Cell Lines
Aннотация
Background: Cancer is a malignant disease characterized by the uncontrolled division of cells, leading to the invasion and destruction of healthy tissues. Copper(II) complexes of 1,10-phenanthroline and its derivatives have demonstrated various biological activities, including anti-tumor, anti-Candida, antimycobacterial, and antimicrobial effects. The aim of the study:To investigate the anti-proliferative effect of Cu(phen)(D-threo)NO₃+ F on the ovarian cancer cell line (A2780) and the colorectal cancer cell line (HT-29). Materials and methods: The copper compound was tested against A2780 and HT-29 cells using the MTT assay at concentrations of 1.0, 2.0, 5.0, 10, 15, and 20 µM, followed by incubation for 24, 48, and 72 hours. IC50 values were determined to assess the compound’s potency. Subsequently, apoptosis induction was evaluated by quantifying caspase-9 protein expression using a human caspase-9 ELISA kit, and DNA fragmentation activity was assessed through agarose gel electrophoresis. Results: The MTT assay revealed that increasing concentrations of the copper compound led to a dose-dependent decrease in cell viability across all incubation periods for both cancer cell lines. The IC₅₀ value for A2780 cells was 1.70±0.26 µM after 72 hours, indicating greater sensitivity compared to HT-29 cells, which exhibited an IC₅₀ of 8.42±0.63 µM. Despite its cytotoxic effects, the compound did not induce apoptosis via the caspase-9 pathway, nor did it promote DNA fragmentation in either cell line, as no significant differences were observed compared to the control group in both assays. Conclusion: Cu(phen)(D-threo)NO₃ + F demonstrated a significant anti-proliferative effect on A2780 and HT-29 cancer cell lines, occurring independently of apoptotic pathways
Ключевые слова: Copper (II) Complex, Anti-Proliferative Activity, Cancer Cell Lines, IC50, Apoptosis Assay
К сожалению, текст статьи доступен только на Английском
Introduction. One of the most potent anti-tumor drugs on the market, cisplatin is frequently used to treat solid tumors such head and neck malignancies, ovarian, lung, and germ cell tumors [1]. It is linked to a number of severe side effects, such as nausea, ototoxicity, and neurotoxicity [2]. Copper has been identified as a more potent and less toxic metal core used in some anticancer drugs. It plays a significant role in endogenous oxidative DNA damage, which is linked to cancer development and aging. As an anti-angiogenic and anticancer agent, a strong copper chelator has demonstrated therapeutic benefits in the treatment of various types of cancer [3].
Copper is a biologically transition metal and it has been extracted for thousands of years; it was known to the Romans as cuprum. The metal is malleable and ductile and an excellent conductor of heat and electricity. Copper is a metal that is usually used to make electrical cables and wires. It contains the element in the +1 and +2 oxidation stress. Copper (I) compounds are mostly white and Copper (II) salts are blue in solution. Numerous proteins and enzymes, including cytochrome oxidase, superoxide dismutase, ascorbate oxidase, and tyrosinase, which are involved in energy metabolism, respiration, and DNA synthesis, depend on it for proper operation. Primarily as a cofactor in several enzymes that promote oxidation-reduction reactions, copper is essential to biological activities. These reactions typically involve copper interacting directly with molecular oxygen, resulting in the formation of free radicals. Given its high reactivity, maintaining proper copper levels is crucial, requiring tightly regulated homeostatic mechanisms. These processes ensure an adequate supply of copper to support essential physiological functions while preventing the toxic effects that can result from excessive copper accumulation [4].
As they can target DNA, copper complexes of 1,10-phenanthroline (Fig. 1) and its derivatives have been used as DNA nucleases in foot printing agents [5]. A number of anticancer drugs known as casiopeinas have been discovered by the modification of the 1,10-phenanthroline copper complex, and one of the complexes has been demonstrated to cause the death of murine leukemia cell lines. Numerous biological processes are disrupted by 1,10-phenanthroline (1,10-phen), 2,20-bipyridine (2,20-bipy), and their substituted derivatives, both in their metal-free form and as ligands coupled to transition metals [6].
The aim of the study. The effects of copper complexes of 1,10-phenanthroline, particularly on their anti-cancer properties, have not been well studied. Thus, investigating Cu(phen)(D-threo)NO3 + F's anti-proliferative impact on colorectal and ovarian cancer cell lines through apoptosis is the main objective of this work.
Materials and Methods
Compound and Cell Lines
Copper (II) complexes of 1,10-phenanthroline and D-threonine associated with NO3 + F, Cu(phen)(D-threo)NO3 + F was supplied by the Department of Chemistry, University of Malaya. Colorectal (HT-29) and ovarian cancer (A2780) cell lines were purchased from the American Type Culture Collection (ATCC), USA.
Chemicals and Reagents
Dulbecco's Modified Eagle Medium (DMEM), Penicillin/Streptomycin antibiotic, Foetal bovine serum (FBS) were purchased from Gibco, Life Technology, UK. Trypsin-EDTA was obtained from ScienCellTM Research Laboratories; ethidium bromide and other standard laboratory chemicals are from Sigma Aldrich, USA and gel powder from Sisco Research Laboratories Pvt. Ltd., Mumbai. MTT reagent was purchased from Calbiochem®, Merck. The kits that I used are DNA extraction kit (iNtRON Biotech., Korea) and caspase-9 kit (Chemicon® Japan).
Cell Seeding
Dulbecco's Modified Eagle Medium (DMEM) was used to cultivate both cancer cell lines on a 96-well plate. 10% fetal bovine serum (FBS) and 1% antibiotics (100U/ml penicillin and 100mg/ml streptomycin) were added to the medium. After cell counting, the cells were prepared based on the concentration that has been set for each well with 1×106 cells/mL. 100µL of the mixture of cells with fresh medium was added into the well respectively. Then, the cells were incubated at 37°C in a humidified atmosphere of 5% CO2 for overnight before the treatment was carried out.
Cell Proliferation Assay and IC50 Value
MTT assay was performed to measure the in vitro anti-proliferation effects of copper compound on cancer cell lines. Briefly, cancer cells were seeded into 96-well plates and left for overnight at 37oC. Each cancer cells were treated with copper (II) complex with different concentrations which are 1µM, 2µM, 5µM, 10µM, 15µM and 20µM and then incubated for 24, 48 and 72 hours. Negative control was treated by using fresh supplemented medium. MTT solution (0.5 mg/mL) was added and incubated for 4 hours. The dark blue formazan crystals formed in intact cells were dissolved in DMSO. The concentration of formazan was reflected in optical density (OD), as measured using a fluorescence microplate reader at 570 and 630 nm [7].
IC50 is the concentration of an inhibitory substance that causes 50% inhibition of cell viability. IC50 for each cancer cell lines can be determined from the dose-response graph as in the result [8].
Human Caspase-9 ELISA Assay
Cancer cells were cultured to a density optimal for apoptosis induction not exceeded 106 cells/mL. The cancer cells were treated with copper compound by using IC50 value. Concurrently, a negative control cell population which is non-induced also incubated at the same density. The sample was prepared for each labelling condition. Four populations have been prepared for Unlabeled, induced and non-induced and FLICA-labeled, induced and non-induced.
The Fluorochrome Inhibitors of Caspases (FLICA) are the basis for Caspase-9 In Situ Assay Kits. A green fluorescence is produced by the carboxyfluorescein-labeled fluoromethyl ketone peptide inhibitor of caspase-9 (FAM-LEHD-FMK) used in this experiment. A reactive cysteine residue on the large subunit of the caspase heterodimer will be covalently bound by this inhibitor. The amount of activated caspase-9 in the cell at the moment the reagent was applied is shown by the green fluorescence signal. Detailed protocol is published elsewhere [9]. 100µL of each cell suspension was placed into each of two wells of a black micro titer plate. The absorbance was read by using an excitation wavelength of 490nm and emission wavelength of 520nm
Measurement of DNA fragmentation
Total DNA was isolated from the pelleted cells using a DNA extraction kit. The DNA was subjected to 2.5% agarose gel electrophoresis in Tris-acetate–EDTA running buffer and stained with 0.5 μg/mL ethidium bromide. DNA stained with ethidium bromide on agarose gels may be seen by transillumination with ultraviolet light and captured on camera with an AlphaImager® HP gel picture system [10].
Statistical analysis
The mean ± standard deviation (SD) was used to express all data. The Statistical Package for Social Science (SPSS) version 24.0 was used to analyze the data using one-way analysis of variance (ANOVA) and then Duncan's multiple range test. Data was considered statistically significant at the value of probability less than 0.05 (P<0.05).
Results.
Anti-proliferation effect of copper (II) complex on cancer cell lines
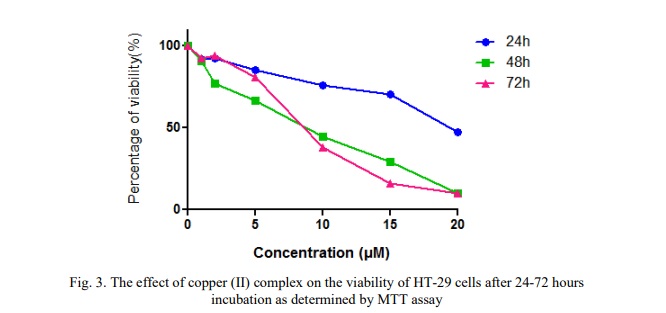
Cu(phen)(D-threo)NO₃ + F was initially incubated with two cancer cell lines, HT-29 and A2780, at various concentrations. The anti-proliferative effects were assessed using the MTT assay. Data obtained for the copper(II) complex following continuous incubation for 24, 48, and 72 hours were used to calculate IC₅₀ values. The compound demonstrated a concentration- and time-dependent anti-proliferative profile in both cell lines.
Table 1 shows the IC50 values of copper (II) complex as tested in A2780 and HT-29 cells at different time incubation. The result indicates that the compound has certain effect on cancer cells and exhibit cytotoxic activity toward the tested human cancer cell lines, especially against A2780 cells. A2780 cells appeared to be more sensitive which showed comparatively lower IC50 values compared to IC50 values of HT-29 cells.

Based on the MTT assay, exposure of A2780 cells to increasing concentrations of the copper(II) complex resulted in a decrease in cell viability, as shown in Figure 2. The dose-response curves indicated a significant reduction (p<0.05) in the percentage of cell viability after A2780 cells were treated with various concentrations of the copper(II) complex for 24 hours. A similar decline in cell viability was observed with increasing concentrations of the complex over 48 and 72 hours of incubation. Cell viability was not significantly altered by the 48- or 72-hour treatments.
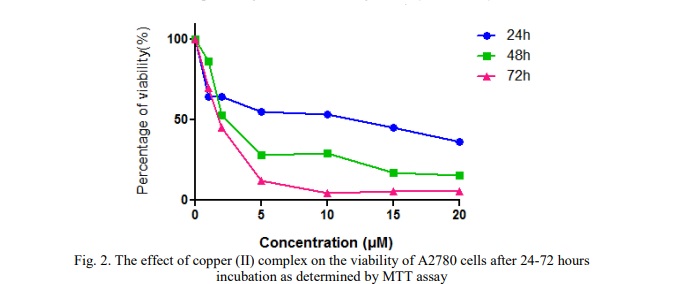
Time-course experiments using different concentration of copper (II) complex up to three days showed that its anti-proliferative activity was already evident after 24 hours of treatment.
Figure 3 shows the dose-response curves, demonstrating a decrease in cell viability in HT-29 cells following treatment with the copper (II) complex at increasing concentrations of 1.0, 2.0, 5.0, 10, 15, and 20 µM over a 24-hour incubation period. The figure also illustrates a reduction in cell viability with increasing concentrations of the complex over 48 and 72 hours of incubation. However, there were no significant differences in cell viability between the 48- and 72-hour treatments at these concentrations.
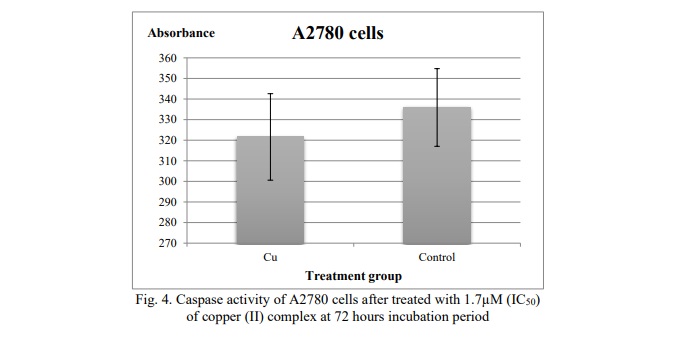
Expression of Caspase-9
The human Caspase-9 ELISA kit was used to measure the amount of caspase activity using a fluorescence microplate reader. The absorbance reading indicates the amount of active caspase-9 present in the cells. Based on Figure 4, the reading of the absorbance for A2780 cells treated with the concentration 1.7µM (IC50) of copper (II) complex was fewer than the reading of the absorbance of control. When compared to the absorbance reading of the negative control, there was no significant change.
DNA fragmentation
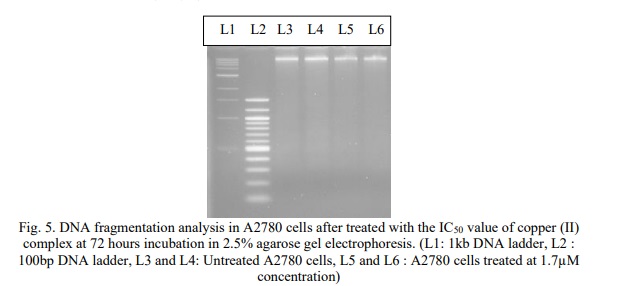
After 72 hours of incubation, cells treated with the IC50 concentration of the copper (II) complex were examined for internucleosomal DNA breakage. Agarose gel electrophoresis was used to isolate the DNA from A2780 cells, which was then stained with ethidium bromide and examined under a UV lamp (Fig. 5). The results showed that DNA from both untreated cells (Lanes 3 and 4) and treated cells (Lanes 5 and 6) remained intact, with no evidence of DNA laddering observed.
Discussion
Anti-proliferative assay (MTT Assay)
The findings of this study demonstrated that increasing concentrations of Cu(phen)(D-threo)NO₃ + F resulted in decreased cell viability in both ovarian (A2780) and colorectal (HT-29) cancer cell lines across 24, 48, and 72 hours of incubation. Initial viability assays revealed that, after 24 hours of treatment, the IC₅₀ value of the copper(II) complex was significantly higher compared to the IC₅₀ values observed at 48 and 72 hours for both cell lines. However, there was no discernible change in the complex's effects at 48 and 72 hours, suggesting that its anti-proliferative action was already evident at 24 hours, when cell viability had dropped approximately 50%. Therefore, the concentrations of Cu(phen)(D-threo)NO₃ + F required to achieve 50% growth inhibition in A2780 and HT-29 cells are higher than the respective IC₅₀ values, which are 1.7 µM and 8.42 µM.
Additionally, the findings showed that this compound was acutely toxic to both human ovarian and colorectal cancer cells. The metal-phen complexes were found to be more potent and to have a cytotoxic effect compared to the most potent simple salt, due to the absence of phen in the simple solvated Cu(II) [11]. The results of the viability study (see Figs. 2, 3 and 4) showed that the Cu(phen)(D-threo)NO₃ + F displayed a concentration- and time-dependent anti-proliferative effect following 24, 48 and 72 hours of exposure. Furthermore, A2780 cells were shown to be most sensitive, with the Cu(phen)(D-threo)NO3 + F compared to HT-29 cells. According to Deegan et al. [12], the copper-phen complex demonstrated the greatest cytotoxic response by inducing a concentration-dependent cytotoxic effect between 3 and 18 times greater than that observed for the metal-based anti-cancer agent, cisplatin. These findings demonstrate that the concentrations of the copper(II) complex required to significantly reduce the viability of cancer cell lines are quite low. For example, 2.8 µM of [Cu(phen)(edda)] was sufficient to decrease the viability of MCF-7 breast cancer cells by 50%, likely due to their higher sensitivity driven by a faster growth rate [12]. Similarly, in this study, the IC₅₀ values for the copper(II) complex were 1.7 µM for A2780 ovarian cancer cells and 8.42 µM for HT-29 colorectal cancer cells, indicating that effective antiproliferative activity occurs at low concentrations.
Human Caspase-9 ELISA Assay
The role of the proteolytic machinery in Cu(phen)(D-threo)NO₃ + F-induced apoptosis was examined using the carboxyfluorescein-labeled fluoromethyl ketone peptide inhibitor of caspase-9 (FAM-LEHD-FMK). The amount of active caspase-9 in the cells at the moment of reagent addition may be directly determined by the green fluorescent signal that results from this inhibitor's covalent binding to active caspase-9. As shown in Figure 4, the number of active caspase-9 molecules in A2780 cells was lower compared to the control, which exhibited higher fluorescence signals. This indicates that treatment with Cu(phen)(D-threo)NO₃ + F did not produce a significant increase in caspase-9 activation relative to untreated cells. Therefore, we can conclude that Cu(phen)(D-threo)NO₃ + F does not induce apoptosis via the caspase-9 pathway. The results are best expressed as the fold increase in caspase activity in apoptotic cells relative to non-induced control cells.
It was found that apoptosome formation in the Cu(phen)(D-threo)NO3 + F could still be induced, but subsequent recruitment and activation of caspase-9 by the complex was defective. These results suggest a novel regulation at the level of caspase-9 activation by the apoptosome complex, and this regulation is repressed in the ovarian cancer cells [13]. Copper complexes were also found to inhibit enzymatic activity and induce cell apoptosis [4]. Surprisingly, our data clearly show that the typical apoptotic pathway induced by Cu(phen)(D-threo)NO3 + F occurs without activation of known caspases. This conclusion is based on the caspase inhibitors, FAM-LEHD-FMK that clearly inhibit caspase-9 have no effect on apoptosis induced by copper Cu(phen)(D-threo)NO3 + F in A2780 cells. Thus, Cu(phen)(D-threo)NO3 + F seems to activate a caspase independent pathway resulting in apoptotic pathway. Cu(phen)(D-threo)NO3+F may activate non-caspase-dependent or alternative pathways of apoptosis, as shown by the comparatively low amount of caspase activation and decreased inhibition by FAM-LEHD-FMK. The mechanism behind the mitochondrial regulation of apoptosis and caspase-independent cell death has been uncovered by recent research. We use the terms caspase-dependent death (apoptosis) and caspase-independent cell death (CICD) to distinguish between these processes, omitting necrosis, which can result from apoptosis failure [14].
Another possible mechanism is that Cu(phen)(D-threo)NO₃ + F does not directly induce apoptosis in A2780 cells. This may be because caspase-9 is either absent or not activated enough in these cells. Without caspase-9, caspase-3 cannot be activated downstream, which stops the apoptosis cascade from starting. The apoptosome complex activates caspase-9, which acts as an upstream initiator caspase and then triggers caspase-3 [15]. Specifically, active caspase-9 cleaves procaspase-3, converting it into its active form. Active caspase-3 is a key executioner caspase responsible for orchestrating the cellular dismantling characteristic of apoptosis [16]. Therefore, the lack of caspase-9 activation may explain why Cu(phen)(D-threo)NO₃ + F does not trigger apoptosis in A2780 cells. It has been established that not all known programmed cell deaths need caspase activation, and more attention is focused on key molecules involved in nonapoptotic or caspase-independent programmed cell death pathways [17]. This study greatly supports our assumption that Cu(phen)(D-threo)NO3+F may cause caspase-independent cell death. Caspase-independent apoptosis has been observed in several cell types [18]. There are numerous reports of programmed cell death in response to activation of death receptors even when caspases are inhibited, suggesting that caspase-independent pathways can be stimulated by death receptors. Examples include death induced by Fas or TNFα that has been characterized as necrotic [19, 20].
More research has shown that programmed cell death (PCD) may happen even when caspases are completely absent, and other noncaspase proteases have been shown to be capable of carrying out PCD [21]. Another assumption that can be made about this condition is that Cu(phen)(D-threo)NO₃⁺F triggered cell death through excessive autophagy. When cells experience excessive autophagy, they die nonapoptotically without caspase activation. It plays a crucial role in cellular remodeling brought on by differentiation, stress, or cytokine-induced damage, and it helps to remove proteins and organelle components [22].
DNA fragmentation
DNA fragmentation is the hallmark of apoptosis and the presence of DNA ladder is a marker for apoptotic cell death. In Figure 5, it has been shown that there was no significant difference between the treated cell and untreated cells (control). The absence of DNA laddering shows that the DNA fragmentation did not occur and we predicted that Cu(phen)(D-threo)NO3+F was not able to induce apoptosis in A2780 cells through DNA fragmentation. Smearing of the DNA in all treatments could result from necrosis caused by the treatments, or from prolonged apoptosis resulting in secondary necrosis. The increasing drug concentration would cause band smearing, suggesting the possible indication of necrotic cell death [23]. Necrosis-like, caspase-independent programmed cell death is characterised by the absence or marginal presence of chromatin condensation and nuclear fragmentation, as well as the disruption of membrane integrity [24].
The cause of the DNA's failure to break down into oligonucleosomal-length pieces remained unclear and requires further research. Nonetheless, several cell lines have been shown to exhibit no DNA laddering during apoptosis [25]. The presence of big DNA fragments in A2780 cells was verified by field inversion gel electrophoresis. Caspase-9 is the upstream caspase in a protease cascade that activates caspase-3, that induces apoptosis. However, this present study has shown that apoptosis was not triggered by caspase-9. Thus, caspase-3 has not been activated by caspase-9 and apoptosis did not occur. Caspase-3 is required for DNA fragmentation and some of the typical morphological changes of cells undergoing apoptosis [26]. Cu(phen)(D-threo)NO3 + F did not induce apoptosis as lack of internucleosomal DNA fragmentation was observed.
Nevertheless, several publications have demonstrated that some cell lines do not display DNA laddering and instead display the primary characteristics of apoptosis when only high molecular weight (HMW) DNA breakage is present. These consist of several epithelial and mesenchymal cell lines, hepatocytes, and lymphocytic leukemia MOLT-4 cells [25]. Based on Figure 5, the band visible on the agarose gel may represent the larger size of the fragmented DNA. Therefore, Cu(phen)(D-threo)NO₃ + F may induce apoptosis in the presence of high-molecular-weight DNA cleavage. Kari et al. [25] reported that apoptosis can also occur in the absence of DNA fragmentation. Since oligonucleosomal DNA fragmentation is not necessary for apoptosis to occur, biochemical tests like DNA ladders should not be utilized to characterize this kind of cell death. Although caspase activation may be necessary for the development of the apoptotic morphology, active inhibition of DNA fragmentation or caspase activation frequently shows that these alterations are not necessary for the execution of the cell death program. However, the diagnosis of apoptosis may benefit from the measurement of caspase activity or DNA fragmentation [27].
Moreover, the absence of normal DNA fragmentation in ovarian cancer cells that have undergone apoptosis suggests that there may be a risk of genetic material being transferred from one tumor cell to another or to nearby healthy cells. Since ovarian carcinoma is a heterogeneous cancer [28], it is plausible that genetic information transmission between cancer cells plays a role in the development of aggressive behavior and resistance to immunotherapeutic and chemotherapy treatment regimens [29]. Furthermore, the activation of caspase-3 is not only relies on the activation of caspase-9 but it also influenced by the other factors. According to Tardito et al. [30], apoptosis is not observed in cell death processes induced by high intracellular copper levels because copper itself inhibits caspase-3 activity. Although there is no direct evidence of a physical interaction between copper and the enzyme, it is plausible that copper interferes with the catalytic cysteine residues within the active site of caspase-3. This inhibitory mechanism supports our findings and leads us to propose that copper may suppress caspase-3 activity, thereby impairing the downstream processes of apoptosis, such as DNA fragmentation.
Conclusion. Cu(phen)(D-threo)NO₃ + F demonstrated potent cytotoxic effects against A2780 and HT-29 cells in a dose- and time-dependent manner. However, apoptosis was not induced via the caspase-9 pathway, and DNA fragmentation was not observed during cell death. While caspase-independent mechanisms, such as mitochondrial-induced apoptosis, cannot be ruled out, their involvement remains uncertain. Therefore, further investigation of alternative pathways for copper-induced apoptosis is required to fully understand the mechanisms underlying its cytotoxic effects.
Financial support
No financial support has been provided for this work.
Conflict of interests
The authors have no conflict of interest to declare.














Список литературы
Список использованной литературы появится позже.