
Перспективы регуляции взаимодействия NK-клеток и клеток трофобласта с помощью ингибиторов циклинзависимых киназ
Aннотация
Актуальность: Анализ взаимосвязи между натуральными киллерами и клетками трофобласта имеет первостепенное значение для понимания процессов репродуктивной иммунологии. Это исследование создает базу для формирования инновационных методов терапевтического воздействия. В данной работе исследуется влияние ингибирования киназ CDK8/19 на функции NК-клеток и их взаимодействия. Данные киназы способствуют развитию и индукции функциональных характеристик клеток иммунной системы. В настоящее время ведутся активные исследования, направленные на разработку стратегий применения ингибиторов киназ в терапии различных заболеваний. Тем не менее, вопрос их использования для модуляции взаимодействий между иммунными клетками в системе мать-плод остается слабо изученным направлением. Это делает актуальным проведение исследований. Цель исследования:Исследование потенциальных эффектов, которые оказывают ИЦК на взаимодействие между NK-клетками и клетками трофобласта в условиях, моделирующих микроокружение при беременности. Материалы и методы:В исследовании использовали клеточные линии NK-92 и JEG-3, культивируемые с цитокинами TNFα, IFNγ, TGFβ1, IL-15, IL-18 и IL-10 в концентрациях, соответствующих их содержанию в биологических жидкостях человека. Экспрессию молекул стресса MICA и активирующих рецепторов NKG2D на NK-клетках оценивали методом проточной цитофлуориметрии после совместного культивирования с клетками трофобласта в присутствии/отсутствии ингибиторов цитокинов, а содержание цитокинов в супернатантах определяли иммуноцитохимическим методом. Результаты:в ходе данного исследования было показано, что ингибитор циклинзависимых киназ (ИЦК) модулирует экспрессию ключевых молекул, таких как активационный рецептор NKG2D и стрессовая молекула MICA, а также влияет на секрецию цитокинов, включая IL-10 и RANTES. Выявлено, что при взаимодействии с клетками трофобласта происходит усиление цитотоксической активности NK-клеток, несмотря на действие ИЦК, что указывает на сохранение ключевых механизмов распознавания мишени и подтверждает важную роль трофобласта в регуляции активности NK-клеток. Кроме того, выявлено дифференцированное влияние провоспалительных цитокинов (IL-18, TNFα, IFNγ) на продукцию IL-10 при предварительной обработке NK-клеток ИЦК, что может свидетельствовать о возможном перекрёсте внутриклеточных сигнальных путей. Заключение:Полученные данные демонстрируют сложную регуляцию взаимодействия NK-клеток и трофобласта и подчеркивают важность дальнейшего изучения возможностей использования ингибиторов CDK8/19 для модуляции иммунного ответа, включая применение в репродуктивной медицине
Введение.NK-клетки являются важным и актуальным объектом исследований в области репродуктивной иммунологии, поскольку они играют ключевую роль в регуляции ранних этапов беременности, включая имплантацию эмбриона и формирование фетально-материнского интерфейса [1, 2]. Их функциональная активность, особенно цитотоксический потенциал и способность к секреции цитокинов, находится под строгим контролем для обеспечения баланса между защитой организма от патогенов и толерантностью к полуаллогенному плоду. Вместе с тем, их участие в развитии репродуктивных заболеваний остается предметом дискуссий и активных научных исследований. Некоторые данные указывают на то, что избыточная цитотоксическая активность NK-клеток может негативно влиять на процесс имплантации и способствовать развитию таких состояний, как привычное невынашивание беременности или преэклампсия [3]. Тем не менее, результаты научных исследований остаются противоречивыми: у некоторых женщин с нарушениями репродуктивной функции отмечено снижение активности естественных киллеров. Это может быть обусловлено нарушением их нормального функционирования, что, в свою очередь, приводит к неспособности обеспечить полноценную поддержку развития плаценты [4, 5]. Таким образом, дальнейшее изучение фенотипических и функциональных характеристик NK-клеток при физиологической и патологической беременности имеет важное значение для понимания механизмов репродуктивных нарушений и разработки целевых иммуномодулирующих терапий.
Наличие NK-клеток в ткани эндометрия создаёт условия для их непосредственного взаимодействия с клетками эмбрионального происхождения, в частности с клетками трофобласта – внешнего слоя бластоцисты [6]. Существует предположение, что трофобластные клетки способны влиять на характеристики и функции материнских NK-клеток как через прямые контактные механизмы, так и путём дистантных взаимодействий благодаря секреции различных биологически-активных веществ [7]. Одним из механизмов опосредованного влияния является секреция цитокинов со стороны клеток трофобласта [8]. Попадая в микроокружение и связываясь с соответствующими рецепторами на поверхности NK-клеток, цитокины активируют внутриклеточные сигнальные каскады, которые могут изменять функциональную активность этих клеток. Среди ключевых цитокинов, которые участвуют этом процессе, можно выделить IL-10, IFNγ и RANTES. Они продуцируются как самими NK-клетками, так и могут влиять как на их собственные характеристики, так и на поведение клеток трофобласта во время беременности. В частности, воздействие RANTES способствует усилению пролиферации и миграции как NK-клеток, так и клеток трофобласта [9, 10]. В то же время, несмотря на способность IL-10 и IFNγ повышать функциональную активность NK-клеток, эти цитокины проявляют антиинвазивное действие в отношении клеток трофобласта [11, 12].
Контактное взаимодействие между различными типами клеток в организме осуществляется посредством сложных лиганд-рецепторных механизмов. Эти механизмы играют ключевую роль в передаче сигналов, которые регулируют активность клеток и их функциональное состояние. Ярким примером такого взаимодействия служит связь между NK-клетками и клетками-мишенями, которая осуществляется через рецептор NKG2D. Этот рецептор экспрессируется на поверхности естественных киллеров и играет важную роль в их функционировании. Данный рецептор способен распознавать стресс-индуцированные молекулы, такие как MICA, которые появляются на поверхности повреждённых, инфицированных или трансформированных клеток. Взаимодействие NKG2D с MICA запускает мощный активационный сигнал, который приводит к реализации цитотоксических функций NK-клеток и последующей гибели клетки-мишени [13]. Однако важно отметить, что MICA может также экспрессироваться на самой поверхности NK-клеток, что играет важную роль в процессе аутокринной регуляции их активности. Подобный механизм играет ключевую роль в предотвращении избыточной активации иммунной системы и способствует поддержанию иммунологической толерантности. Это особенно важно в физиологических состояниях, таких как беременность, когда требуется тонкий баланс между эффективной защитой от патогенов и предотвращением иммунного ответа против полугенного плода [14]. Несмотря на значимость этих механизмов, состояние стресса у NK-клеток до сих пор остаётся недостаточно изученным явлением. Не до конца понятно, какие именно факторы могут вызывать стрессовые реакции в этих клетках, а также как это влияет на их функциональные свойства и взаимодействие с другими клеточными элементами. Кроме того, остаётся открытым вопрос о роли клеточных компонентов микроокружения в контексте физиологической беременности — в частности, клеток трофобласта, которые потенциально могут инициировать или модулировать стрессовые реакции в NK-клетках.
Существуют значительные пробелы в понимании молекулярных механизмов, управляющих функциональной активностью NK-клеток и их взаимодействием с клетками трофобласта, что обуславливает необходимость поиска потенциальных регуляторов этих процессов для создания новых терапевтических стратегий.
Циклинзависимые киназы (CDK) представляют собой важную группу ферментов, участвующих в регуляции клеточного цикла и процессов транскрипции, обеспечивая своевременное развитие клеточных событий через взаимодействие с регуляторными субъединицами – циклинами [15]. Многие представители этого семейства киназ играют ключевые роли в передаче сигналов от внешних стимулов к ядру клетки, включая реакцию на действие различных цитокинов и других сигнальных молекул [16, 17]. Среди них особое внимание привлекают CDK8 и её близкий гомолог CDK19, являющиеся частью медиаторного комплекса, который взаимодействует с РНК-полимеразой II и участвует в регуляции инициации транскрипции [15].
В составе медиаторного комплекса CDK8/19 способны связываться с различными факторами транскрипции, обладающими специфичностью к определённым последовательностям ДНК, тем самым оказывая влияние на экспрессию генов, вовлечённых в ключевые клеточные процессы и сигнальные пути [18]. Благодаря своему участию в регуляции транскрипции и клеточной активности, CDK8/19 рассматриваются как потенциальные терапевтические мишени, особенно в контексте заболеваний, связанных с дисрегуляцией иммунного ответа и воспалительных реакций.
В настоящее время активно разрабатываются и изучаются ингибиторы CDK8/19 (ИЦК), которые демонстрируют высокую эффективность в модуляции транскрипционной активности и могут быть использованы в качестве перспективных терапевтических агентов при различных заболеваниях, включая онкологические и аутоиммунные [19, 20, 21]. Несмотря на явные перспективы данных соединений, до настоящего времени не проводилось исследований, направленных на оценку влияния этих соединений на функциональную активность NK-клеток и их взаимодействие с клетками трофобласта — процесс, имеющий критическое значение для успешного развития беременности. Такие исследования могли бы расширить понимание механизмов иммунной регуляции в условиях физиологической и патологической беременности и открыть новые возможности для создания целевых иммуномодулирующих препаратов.
Цель исследования. Исследование потенциальных эффектов, которые оказывают ИЦК на взаимодействие между NK-клетками и клетками трофобласта в условиях, моделирующих микроокружение при беременности.
Материалы и методы исследования
Клетки
В исследовании были использованы хорошо охарактеризованные и широко применяемые в научной практике клеточные линии: NK-92 и JEG-3. Линия NK-92 была получена в 1992 году из периферической крови мужчины, страдающего быстро прогрессирующей неходжкинской лимфомой, и представляет собой активированные естественные киллеры (NK-клетки), обладающие выраженной цитотоксической активностью. Эта клеточная модель часто используется для изучения функциональных свойств NK-клеток, поскольку сохраняет ключевые характеристики первичных NK-клеток, включая экспрессию важнейших рецепторов и способность к лизису мишеней [22].
Для изучения взаимодействия с NK-клетками в качестве клеток-мишеней была выбрана трофобластоподобная клеточная линия JEG-3, выделенная в 1974 году из штамма Эрвина-Тернера опухоли Вудса. JEG-3 обладает рядом свойств, характерных для трофобласта, включая секрецию гормонов, таких как хорионический гонадотропин, а также экспрессию различных мембранных молекул, участвующих в иммунном взаимодействии, что делает её удобной моделью для исследования фетально-материнских взаимоотношений на клеточном уровне [22, 23].
Культивирование обеих клеточных линий проводилось в строгом соответствии с рекомендациями, установленными поставщиком — коллекцией Американской типовой культуры (ATCC, США). Для поддержания жизнеспособности и пролиферативной активности клеток использовались соответствующие питательные среды, дополненные необходимыми ростовыми факторами, антибиотиками и сывороткой животных. Все эксперименты проводились с соблюдением условий асептики и контролируемого температурного режима, что обеспечивало воспроизводимость результатов и корректность интерпретации данных. Использование указанных клеточных моделей позволило создать адекватную in vitro систему для изучения механизмов взаимодействия NK-клеток и клеток трофобласта, а также оценки влияния различных модуляторов на их функциональную активность [22, 23].
Индукторы
Использовали цитокины TNFα (50 U/ml), IFNγ (1000 U/ml), TGFβ1 (5 ng/ml), IL-15 (10 ng/ml), IL-18 (10 ng/ml), IL-10 (10 ng/ml), ("RnD", США)).
Указанные концентрации цитокинов соответствуют содержанию в биологических жидкостях человека, в том числе в зоне маточно-плацентарного контакта [24, 25].
Оценка экспрессии MICA и NKG2DNK-клетками в присутствии клеток трофобласта и ИЦК
Клетки трофобласта линии JEG-3 высевали в культуральные флаконы для адгезивных клеточных культур площадью 25 см² (BD, США) в концентрации 200 000 клеток на 1 мл (общее количество – 1 000 000 клеток). По истечении 24 часов для окрашивания 1,5×10⁶ клеток линии NK-92 применяли сукцинимидиловый эфир карбоксифлуоресцеина (СFSE, сarboxyfluorescein succinimidyl ester) («Sigma-Aldrich», CША), соблюдая рекомендации, указанные производителем [7]. Для этого раствор CFSE добавляли во флаконы с клетками линии NK-92, после чего инкубировали при температуре 37°C, 5% CO₂ в течение 10 минут. Окраску останавливали, помещая на холод, затем отмывали холодным α-МЕМ («Биолот», Россия), смешанный с бычьим сывороточным альбумином, доведенным до концентрации 0,5%. После обработки CFSE клетки линии NK-92 разводили в 5 мл полной ростовой среды для NK-клеток до концентрации 200 000 клеток на 1 мл и переносили во флаконы с клетками трофобласта линии JEG-3. Для обозначения варианта совместного культивирования клеток линий NK-92 и JEG-3 в тексте использовали термин «сокультура». В качестве контроля для оценки изменений фенотипа применяли интактные клетки линии NK-92, окрашенные CFSE и культивируемые в полной среде для NK-клеток, а также интактные клетки линии JEG-3, прокультивированные в полной среде для NK-клеток. Далее во флаконы с клетками линии NK-92, JEG-3, а также во флаконы с сокультурой добавляли цитокин TGFβ в концентрации 5 нг/мл. За 60 минут до начала эксперимента часть клеток культивировали в присутствии ИЦК (Кортистатин А) в дозе, не оказывающей влияния на жизнеспособность, после чего к ним также приливали цитокин TGFβ. После этого клетки инкубировали в течение 24 часов. Затем производили дезинтеграцию образовавшегося монослоя с помощью культурального скребка, содержимое переносили в пробирки и центрифугировали (300g, 10 мин.), обрабатывали раствором «Fc-block» для предотвращения неспецифического связывания антител (Miltenyi Biotec, Испания), придерживаясь инструкций производителя [7]. Затем клетки подвергали обработке моноклональными антителами к NKG2D и MICA (BD, США) в соответствии с инструкциями производителя. Для контроля неспецифического связывания антител применяли изотипические антитела (BD, США). После инкубации при температуре 4°C в течение 20 минут проводили оценку фенотипа и интенсивности флуоресценции клеток с использованием проточного цитофлуориметра FacsCantoII (BD, США).
Оценка концентрации цитокинов в супернатантах, полученных в результате инкубации клеток линии NK-92.
Клетки переносили в культуральные флаконы объемом 25 см² («Sarstedt», Германия) по 900 000 клеток в 3 мл полной ростовой среды. Во все флаконы добавляли IL-2 (500 Ед/мл) («Биотех», Россия). Часть клеток культивировали без добавления индукторов, часть культивировали в присутствии ИЦК. Спустя 60 минут к клеткам в указанных ранее концентрациях добавляли цитокины TNFα, IFNγ, TGFβ1, IL-15, IL-18 или IL-10, при этом часть клеток оставляли без индукторов в качестве контрольных образцов. Через 72 часа для продолжения стимуляции к клеткам вносили цитокины без или с добавлением ИЦК в 1,5 мл среды. Спустя 24 часа содержимое флаконов центрифугировали в течение 5 минут при 2500g, супернатанты замораживали при температуре -20ºС. Для определения уровня IL-10, RANTES и IFNγ применяли тест-наборы (BD, США), выполняя процедуру согласно предоставленным производителем указаниям. Измерения проводили с использованием проточного цитофлуориметра FACSCanto II («BD», США).
Статистический анализ полученных данных.
Каждый эксперимент включал четыре независимых биологических повтора, при этом в рамках каждого биологического повтора выполнялось по два технических повтора для каждого исследуемого условия культивирования. Статистическая обработка экспериментальных данных осуществлялась с использованием программного обеспечения GraphPad Prism 8. Предварительная проверка на нормальность распределения выборок проводилась с применением критерия Шапиро–Уилка. Далее использовали непараметрический аналог t-критерия Стьюдента – тест Манна-Уитни. Статистически значимыми признавали различия при p <0,05.
Анaлиз экспрессииNК-клетками рецептора NKG2D и его лигандаMICАбез и в присутствии TGFβ и ИЦК
Установлено, что клетки линии NK-92 спонтанно экспрессируют на совей поверхности рецептор NKG2D (99,74 {99,58; 99,89}) и стрессорную молекулу MICA (32,75 {31,87; 41,81}) (Рис. 1А) (данные об экспрессии молекул отражены в таблице 1 «Уровень экспрессии рецептора NKG2D и стресс-молекулы MICA на естественных киллерах (NK-клетки)», отражающей медиану {25% перцентиль; 75% перцентиль}), что согласуется с полученными нами ранее данными по фенотипу для данной клеточной линии [14]. Количество клеток линии NK-92, экспрессирующих активирующий рецептор NKG2D на своей поверхности, оставалось неизменным как при моно и сокультивировании (Рис. 1А). При этом было отмечено увеличение интенсивности экспрессии рецептора NKG2D клетками линии NK-92 после их совместного культивирования с клетками трофобласта линии JEG-3 по сравнению с интактными клетками NK-92 (Рис. 1В). При этом, интенсивность экспрессии рецептора NKG2D NK-клетками не менялась ни при воздействии индукторов в моно-, ни в сокультуре.
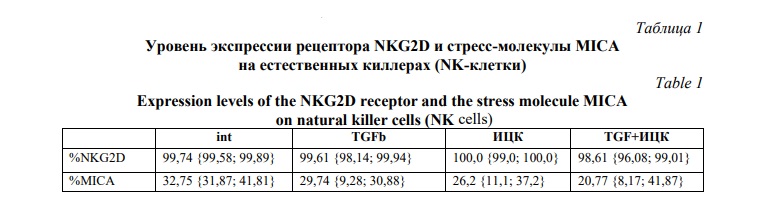
При культивировании клеток линии NK-92 в присутствии цитокина TGFβ наблюдалось снижение числа клеток, демонстрирующих экспрессию стресс-ассоциированной молекулы MICA, в сравнении с контролем без добавления цитокинов (Рис. 1С). После совместного культивирования с клетками JEG-3 также было зафиксировано уменьшение количества клеток, экспрессирующих молекулы MICA, по сравнению с монокультурой NK-клеток (Рис. 1С). Другие протестированные индукторы не оказали значимого влияния на уровень экспрессии MICA ни в моно-, ни в сокультуре.
Интенсивность флуоресценции молекулы MICA на клетках линии NK-92 увеличивалась после их совместного культивирования с клетками трофобласта линии JEG-3 по сравнению с исходным уровнем (Рис. 1D). Однако обработка ингибиторами ИЦК приводила к снижению данного показателя (Рис. 1D). Другие индукторы, использованные в работе, не влияли на интенсивность экспрессии стресс-ассоциированной молекулы MICA клетками линии NK-92.
Анализ секретома NK-клеток в присутствии ИЦК и цитокинов
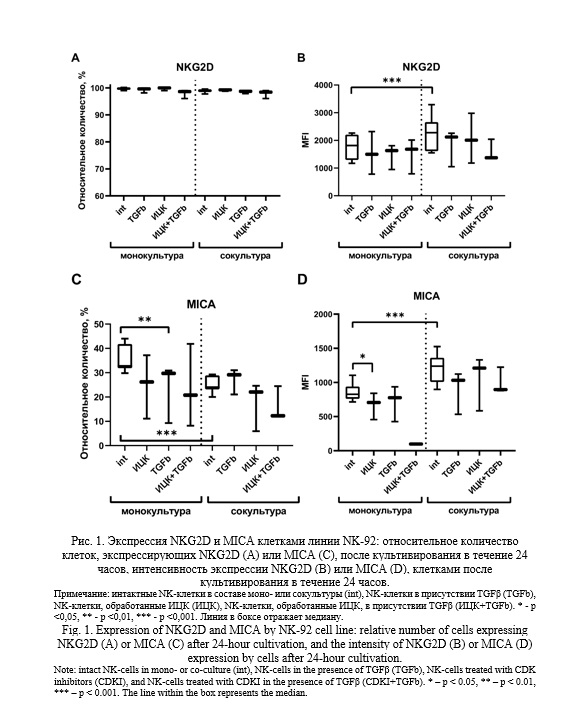
Выявлено, что клетки линии NK-92 спонтанно продуцируют цитокины IFNγ, RANTES, IL-10, что согласуется с данными литературы [26, 27].
Уровень цитокина IFNγ в супернатанте снизился после культивирования клеток, предварительно подвергнутых воздействию ИЦК, в присутствии IL-10, по сравнению с его содержанием в средах после культивирования клеток линии NK-92, необработанных ИЦК, и прокультивированных с цитокином (Рис. 2А). При иных условиях культивирования изменений концентрации цитокина в супернатанте обнаружено не было. Концентрация RANTES была выше в проанализированных кондиционированных средах, полученных от культивирования клеток линии NK-92 в присутствии ИЦК по сравнению с необработанными клетками, однако при добавлении цитокинов TGFβ, IL-10 или IL-18 этот эффект ИЦК отменялся, количество RANTES снижалось. Также концентрация RANTES в супернатантах понижалась при добавлении TGFβ к клеткам линии NK-92 относительно концентрация RANTES в супернатантах от интактных клеток. В отличие от TGFβ, другие цитокины не оказывали подобного эффекта на уровень RANTES.
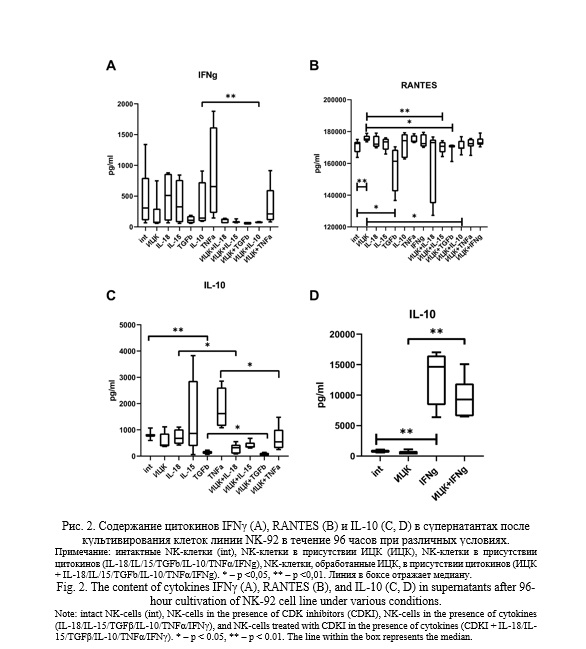
Концентрация цитокина IL-10 в супернатантах снижалась после культивирования клеток линии NK-92 в присутствии TGFβ относительно необработанных клеток, однако предварительная обработка клеток ИЦК приводила к еще большему снижению содержания цитокина в системе. Кроме того, предварительная обработка клеток ИЦК приводила к снижению содержания IL-10 при культивировании с цитокинами IL-18 или TNFα по сравнению с содержанием белка в среде, полученной от культивирования в присутствии только цитокинов. Культивирование клеток линии NK-92, предварительно обработанных ИЦК, в присутствии цитокина IFNγ вызывало повышение уровня белка в супернатантах после культивирования по сравнению с его содержанием в супернатантах, полученных при культивировании клеток без добавления цитокина.
Заключение. Взаимодействие NK-клеток с клетками трофобласта представляет собой ключевое направление исследований в современной репродуктивной иммунологии и требует дальнейшего изучения с целью поиска возможных способов его регуляции, которые могут лечь в основу разработки новых терапевтических подходов для профилактики и лечения заболеваний, возникающих при беременности. Одной из потенциальных мишеней для фармакологического воздействия могут являться циклинзависимые киназы, ингибирование которых способно изменять экспрессию генов в клетках, модулируя их фенотипические характеристики и секретом, что, в свою очередь, влияет на функциональную активность клеток [28]. В рамках настоящего исследования изучалось влияние селективного ингибирования CDK8 и CDK19 на взаимодействие NK-клеток с клетками трофобласта — процесс, играющий решающую роль в имплантации эмбриона и последующем развитии беременности. Полученные данные могут способствовать углублению понимания молекулярных механизмов, регулирующих фетально-материнские взаимодействия, и открыть новые перспективы для иммуномодулирующей терапии при репродуктивных заболеваниях.
На первом этапе исследования оценивали экспрессию активационного рецептора NKG2D, а также маркера стресса MICA NK-клетками линии NK-92. При культивировании в присутствии цитокина TGFβ количество NK-клеток, экспрессирующих MICA на совей поверхности снижалось.
Кроме того, было установлено, что интенсивность экспрессии молекулы MICA на поверхности клеток линии NK-92 снижалась после культивирования с ИЦК. Это может свидетельствовать о снижении уровня клеточного стресса и уменьшении риска их распознавания и лизиса другими иммунными клетками, что имеет значение для регуляции активности NK-клеток в физиологических и патологических условиях.
Однако в сокультуре с клетками трофобласта описанные эффекты отменялись. Поскольку интенсивность экспрессии NKG2D и MICA увеличена в сокультуре с клетками трофобласта по сравнению с монокультурой, вероятно в присутствии ИЦК или TGFβ происходят сходные события, экспрессия рецептора NKG2D и стрессорной молекулы MICA NK-клетками компенсируется, что выражается в отмене ингибирующих эффектов ИЦК или TGFβ. Одновременно с этим повышение уровня экспрессии рецептора NKG2D на поверхности NK-клеток в условиях сокультивирования свидетельствует об активации их цитотоксических свойств при контакте с клетками трофобласта [29, 30]. Повышение уровня экспрессии NK-клетками MICA может свидетельствовать об индуцированном усиленным надзором за ними со стороны киллерных клеток иммунной системы, приводя к сокращению численности популяции NK-клеток из-за их элиминации [31, 32]. Полученные данные могут отражать роль клеток трофобласта в регуляции количества и функциональной активности NK-клеток матки при беременности.
Поскольку цитокиновое и клеточное микроокружение способствует приобретению характеристик децидуальных NK-клеток в течение длительного времени [33, 34], для изучения секретома NK-клеток нами было принято решение о пролонгированном культивировании в присутствии ИЦК и факторов, характерных для беременности (время культивирования составило 96 часов).
Ранее мы обнаружили цитокины IFNγ, RANTES и IL-10 в супернатантах, полученных после культивирования интактных клеток линии NK-92 [35]. Данные цитокины также характерны для NK-клеток, наши данные согласуются с данными литературы [26, 27]. В данной работе исследовали изменения цитокинового профиля NK-клеток под воздействием ИЦК, а также цитокинов, типичных для микроокружения во время беременности.
В ходе предыдущих исследований нами были выявлены цитокины IFNγ, RANTES и IL-10 в супернатантах, полученных после культивирования интактных клеток линии NK-92 [35]. Стоит отметить, что данные цитокины являются характерными маркерами для NK-клеток, что подтверждается многочисленными литературными источниками. Наши результаты полностью согласуются с ранее опубликованными данными [26, 27]. В данной работе исследовали изменяли цитокиновый профиль NK-клеток под воздействием ИЦК, включая цитокины, характерные для микроокружения во время беременности. Это дополнительно подчеркивает достоверность и корреляцию наших данных с современными представлениями о функциональной активности NK-клеток.
В присутствии TGFβ в супернатантах выявлено пониженное содержание IL-10 по сравнению с интактными клетками, однако культивирование в присутствии ИЦК вызывало еще большее понижение содержания IL-10 в средах. IL-10 может выступать в качестве стимулятора эффекторных функций NK-клеток [36]. В то же время, этот цитокин способен подавлять инвазивный потенциал клеток трофобласта [11]. Таким образом, снижение продукции IL-10 под действием TGFβ может служить одним из механизмов регуляции активности NK-клеток со стороны клеток трофобласта и других компонентов микроокружения.
Продукция NK-клетками IL-10 не менялась в ответ на добавление ИЦК, IL-18 или TNFα по отдельности. Однако культивирование в комбинации ИЦК+IL-18 или ИЦК+TNFα приводило к снижению секреции IL-10 NK-клетками. Культивирование предварительно обработанных ИЦК клеток в присутствии IFNγ вызывало увеличение содержания IL-10 в супернатантах, относительно его уровня в средах, полученных от культивирования клеток, обработанных только ИЦК. В литературе имеются данные об участии CDK8/19 в ингибировании процесса синтеза IL-10 [37]. Таким образом, цитокины IL-18, TNFα и IFNγ в условиях преинкубации NK-клеток с ИЦК по-разному проявляют действие в отношении продукции IL-10: IL-18 и TNFα снижают, а IFNγ повышает секрецию IL-10 клетками линии NK-92. Нам не удалось найти в литературе данных о роли CDK8/19 или их ингибиторов в индукции клеточного ответа на действие IL-18 или TNFα. Вероятно, в этом случае возможен перекрест внутриклеточных сигнальных путей цитокинов и ИЦК, что приводит к появлению нового эффекта. Однако это предположение требует дополнительной проверки.
Обработка NK-клеток ИЦК стимулировала продукцию ими RANTES. Показано, что для начала синтеза мРНК RANTES требуется активация белков, которые регулируют транскрипцию интерферонов (от англ. interferon regulatory factors - IRF) и NF-kB [38]. В то же время ИЦК могут подавлять элонгацию NF-kB-индуцированной транскрипции в клетках линий, отличных от NK-92 [39]. Таким образом, ИЦК потенциально могут регулировать функции NK-клеток за счет увеличения синтеза RANTES. Однако в присутствии цитокинов IL-15 или TGFβ отмечалось снижение продукции RANTES клетками линии NK-92.
В рамках исследования была проанализирована возможность регуляции как контактных, так и дистантных взаимодействий между клетками трофобласта и NK-клетками, опосредованных цитокинами, с использованием ингибиторов циклинзависимых киназ 8/19 (ИЦК). Результаты показали, что клетки трофобласта могут усиливать цитотоксическую активность NK-клеток путем повышения экспрессии активирующего рецептора NKG2D на поверхности естественных киллеров. Кроме того, установлено, что прямой контакт с клетками трофобласта стимулирует экспрессию стресс-ассоциированной молекулы MICA на мембране NK-клеток. Это может играть важную роль в регуляции их активности, обеспечивая обратную связь со стороны других иммунных клеток через механизм аутокринного взаимодействия.
Культивирование клеток в присутствии ИЦК не приводило к блокированию данных эффектов, вызванных клетками трофобласта, что указывает на сохранение ключевых механизмов взаимодействия NK-клеток с мишенью при воздействии этого фактора. Полученные данные имеют важное значение для понимания влияния ИЦК на иммунные клетки и открывают перспективы для их применения в клинической практике, особенно в онкологии, где модуляция активности естественных киллеров играет ключевую роль в обеспечении противоопухолевого иммунного ответа. Результаты могут способствовать разработке более эффективных и безопасных терапевтических стратегий.
Анализ влияния цитокинов IL-18, TNFα и IFNγ на NK-клетки в присутствии ИЦК выявил различия в их воздействии на продукцию противовоспалительного цитокина IL-10. Это указывает на возможное взаимодействие внутриклеточных сигнальных путей, модулируемых ингибиторами CDK8/19. Полученные данные подчеркивают важность учёта таких перекрёстных взаимодействий при разработке стратегий регуляции активности NK-клеток и их взаимодействия с клетками трофобласта в условиях беременности.
Информация о финансировании
Поддержано ФНИ №1024032800230-9-3.2.2, «Универсальные и специфические механизмы реализации и нарушений репродуктивной функции в семье», рук. д.м.н., профессор, член-корреспондент РАН Коган И.Ю.














Список литературы