
Влияние материнской депривации на уровень внеклеточного дофамина при моделировании состояния азарта
Aннотация
Актуальность:Отлучение от матери (MD) в раннем онтогенезе вызывает у животных стойкие изменения эмоциональных реакций и формирования зависимости от психотропных средств. В настоящем исследовании мы моделировали аддиктивное состояние азарта и риска дофаминовой природы, которое лежит в основе формирования игрового расстройства поведения. Цель исследования:Изучить дофаминовые механизмы аддиктивного состояния у крыс после MD с помощью анализа высвобождения дофамина во внеклеточное пространство прилежащего ядра в ответ на электрическое раздражение зоны самостимуляции вентральной области покрышки (VTA) в модели подкрепления, приближенной к ситуации азарта и риска азартных игр. Материалы и методы:Крысят со 2-го по 12-й день постнатального периода помещали в индивидуальные емкости на 180 мин в течение 10 последовательных дней. У взрослых крыс вживляли электроды в VTA и прилежащее ядро. Животных обучали реакции самостимуляции VTA в фиксированном режиме FR3 в камере Скиннера с гладким полом, полупрозрачными стенами и включением лампочки при нажатии педали. Регистрацию высвобождения дофамина осуществляли в прилежащем ядре методом циклической вольтамперометрии с быстрым сканированием в ответ на включение лампочки и одновременное раздражение VTA в камере Скиннера между сериями самостимуляции. Животных также обучали реакции самостимуляции в режиме переменного соотношения VR6 и тестировали в камере Скиннера с решетчатым полом, стенами белого цвета и включением звука при нажатии педали. Регистрацию высвобождения дофамина осуществляли аналогичным образом. Результаты:У контрольных крыс уровень внеклеточного дофамина в ответ на стимуляцию VTA при включении условных сигналов (лампочки или звука) в камере Скиннера повышался, однако статистический анализ не показал значимых отличий при использовании режимов FR3 и VR6. У крыс MD уровень внеклеточного дофамина в ответ на стимуляцию VTA при подаче условного сигнала в камере Скиннера повышался в большей степени при использовании режима VR6 по сравнению с FR3. Заключение:Таким образом, стресс материнской депривации в раннем онтогенезе вызывает повышение высвобождения дофамина во внеклеточную среду прилежащего ядра в ответ на стимуляцию зоны положительного подкрепления VTA, которая сопровождается подачей условного сигнала аддиктивного состояния самостимуляции в режиме переменного соотношения, приближенной к ситуации азарта и риска
Ключевые слова: материнская депривация, внеклеточный дофамин, самостимуляция, режим переменного соотношения
Введение. Действие стрессоров в раннем возрасте повышает вероятность формирования психических и аффективных расстройств, употребление алкоголя и психоактивных средств [1]. Снижение социальных взаимодействий в онтогенезе, отлучение от матери в раннем возрасте вызывают нарушения поведения и эмоционально-мотивационной сферы [2], тревожные состояния и снижение настроения [3, 4]. Материнская депривация (MD) в раннем онтогенезе приводит к отсроченным последствиям в виде нарушений аддиктивного поведения, зависимости от алкоголя и психоактивных веществ [5, 6]. MD проводится у грызунов при отделении матери от детенышей (от 15 мин до 6 час в день) в течение первых 12 дней после рождения. Ранее мы показали повышение проявления импульсивности, компульсивности и поведения риска у животных MD. Исследовали проявления импульсивности в модели игровой зависимости в модификации теста Iowa Gambling Task. Компульсивный компонент в поведении тестировали в marble тесте закапывания шариков [7]. В проведенных нами исследованиях MD вызывало у крыс также приступообразное (компульсивное) переедание, которое относится к проявлению пищевой зависимости. Оно наблюдалось после 1 мес подачи высокоуглеводной пищи в прерывистом режиме. При этом животные съедали стандартную пищу в том же объеме [8]. В настоящем исследовании мы моделировали аддиктивное состояние при стимуляции системы внутреннего подкрепления при использовании режима, приближенного к ситуации азарта и риска азартных игр.
В работе регистрировали фазические повышения внеклеточного уровня дофамина у животных MD в прилежащем ядре (NAc) в ответ на стимуляцию зоны положительного подкрепления вентральной тегментальной области (VTA). NAc и VTA, структуры дофаминовой системы мозга, играют важную роль в формировании механизмов подкрепления и регуляции стресса в онтогенезе. Система дофамина развивается на протяжении всего периода до полового созревания, а воздействие стрессовых и аверсивных раздражителей может ее нарушить. Во время постнатального развития наблюдаются изменения плотности рецепторов дофамина, содержания тирозингидроксилазы, базального дофамина и экспрессии его рецепторов [9].
Известно, что у крыс, подвергшихся MD, наблюдаются повышенный уровень дофамина в полосатом теле и гиперактивность дофаминергической системы в ответ на стресс во взрослом возрасте [10]. Показано, что у крыс, которые были отделены от матери в раннем возрасте, снижалось содержание транспортеров обратного захвата дофамина, в вентральном стриатуме [11]. Разлучение с матерью снижало внеклеточное содержание транспортера дофамина и его афинности к медиатору, увеличивая время обратного захвата из внеклеточной жидкости [12, 13]. Это может быть связано с более сильным и стойким увеличением синаптического дофамина в ответ на различные стрессоры. В то же время в доступной литературе исследований высвобождения дофамина в ответ на стимуляцию зоны подкрепления у крыс MD показано не было. Результаты исследований на грызунах согласуются с идеей о том, что материнская забота может изменить развитие систем, которые регулируют реакцию на стресс [14]. У людей, которые сообщали о недостаточном родительском уходе в детском возрасте, наблюдалось значительное высвобождение дофамина в вентральном стриатуме, на что указывало снижение потенциала связывания раклоприда в состоянии стресса [15].
Цель исследования. Изучить дофаминовые механизмы аддиктивного состояния у крыс MD с помощью анализа высвобождения дофамина во внеклеточное пространство NAc в ответ на электрическое раздражение зоны самостимуляции VTA в модели подкрепления, приближенной к ситуации азарта и риска азартных игр.
Материал и методы исследования
Выбор животных для эксперимента. Опыты сделаны на 12 крысах самцах породы Вистар весом 240-300 г. До начала экспериментов из питомника «Рапполово» (Ленинградская область) были получены беременные самки. Крыс содержали в виварии при стандартных условиях разведения, когда животные могли свободно получать воду и пищу. Поддерживали освещение с 8.00 до 20.00 и температуру окружающего воздуха – 22±2°C. Эксперименты проведены на основе этических принципов, отмеченных в Директиве Европейского парламента и Совета Европейского Союза 2010/63/EC от 22.09.2010 г. и решении комиссии по биоэтике ФГБНУ «Институт экспериментальной медицины».
Модель МД – материнской депривации. После рождения животных отделяли от матерей с 2-го по 12-й день жизни. Они находились в индивидуальных емкостях в течение 3х часов, десяти дней и без зрительного контакта с матерью. После окончания периода MD и срока молочного вскармливания животных содержали в стандартных клетках по 5 особей. В эксперимент поступали половозрелые самцы в возрасте 90-100 дней и весом 200-250 г.
Операция по вживлению электродов. В вентральную тегментальную область (VTA) с помощью стереотаксиса вживляли электрод для раздражения (стальной электрод 0,2 мм в диаметре). Координаты не изолированного кончика электрода относительно шва брегмы: АР = -5,3 мм, SD = 0,8 мм, Н = 8,2 мм [16]. Для отведения уровня DA в NAc вживляли углеродный электрод в стеклянной изоляции (длина не изолированного кончика электрода 100 мкм, диаметром 7 мкм). Электрод для отведения вживляли по координатам: AP = +2,0 мм (от шва брегмы); SD = 1,2 мм; Н=7,3 мм от поверхности черепа после удаления надкостницы [16]. Имплантировали дополнительно электрод сравнения толщиной 3 мм из Ag/AgCl согласно координатам: АР=+5,5мм (от шва брегмы); SD=0. В условиях стереотаксической операции устанавливали наличие внеклеточного дофамина, затем электроды фиксировали, применяя акрил с УФ-отверждением [17]. После оперативного вмешательства крысы содержались в индивидуальных клетках.
Опыты по исследованию реакции самораздражения проведены на 2х группах: 1. Контрольная группа животных (СG; n=9) и 2. Группа животных после материнской депривации (MD; n=9). В процессе проведения хирургических операций по вживлению электродов 2 животных (1 СG и 1 MD) не выжили. При тестировании реакции самостимуляции 4 животных (2 СG и 2 MD) не удовлетворяли критериям реакции, нажимая педаль менее 5 нажатий/мин и были изъяты из эксперимента. Использовали 2 камеры Скиннера с различными сенсорными характеристиками. В камерах Скиннера фиксировали количество нажатий на педаль и пороговые значения тока (в мкА) за 5 мин опыта. В опытах сначала применяли фиксированный режим подкрепления FR3 (каждое 3е нажатие педали подкрепляли стимуляцией VTA, длительность прямоугольного импульса 1мс с частотой 100 Гц в течение 0,5 с) в камере с гладким полом и полупрозрачными стенами из пластика при включении лампочки 60лк в течение 0,5 с после нажатия педали с подачей подкрепления. Порог стимуляции определяли при возрастающих значениях тока с шагом по 2 мкА до появления реакций нажатия педали. После этого силу тока увеличивали на 50% и затем ее снижали пошагово до отказа от нажатий педали [18].
После стабилизации реакции самостимуляции в режиме FR3 регистрировали высвобождение дофамина. Затем животных обучали реакции самостимуляции в режиме переменного соотношения VR6 и тестировали в камере Скиннера с решетчатым полом и стенами белого цвета при включении звука после нажатия педали (90дБ, 800Гц в течение 0,5с) [19] во время нажатия педали, которое сопровождалось в данном режиме включением электрического тока. При использовании режима переменного соотношения VR6 животное не может определить, какое из шести последовательных нажатий будет сопровождаться подачей электрического тока. Показано, что данный режим наиболее приближен к ситуации азарта и риска азартных игр у человека [20]. Во время непредсказуемого подкрепления животные демонстрировали высокую скорость реакции даже после серии отсутствия подкрепления [21]. При тестировании реакции самостимуляции использовали силу тока, на 50% выше пороговой для самостимуляции.
Определение высвобождения DA производили в свободном поведении при стимуляции VTA [18] в жилой клетке или в камерах Скиннера между сериями самостимуляции по 5 мин каждая, оценивая DA в прилежащем ядре методом циклической вольтамперометрии с быстрым сканированием (FSCV). Навязанная электростимуляция зоны самораздражения VTA сопровождалась включением лампочки в камере (60лк, длительностью 0,5 с) или звука (90дБ, 800Гц в течение 0,5с), и производили запись вольтамерограммы. Стимуляцию VTA производили при подаче пачки импульсов (сила тока, на 50% выше пороговой для самостимуляции, длительность импульса 1 мс с частотой 100 Гц в течение 0,5 сек).
При определении уровня дофамина потенциал удержания составлял 0,4В, а продолжительность сканирования ‒ 9 мс. Через каждые 100 мс подавали сканирующий импульс. Величина анодного предела составляла +1,3В. При обработке материала использовали веб-приложение Analysis Kid, которое позволяет визуализировать, калибровать, и фильтровать нейрохимические сигналы [18].
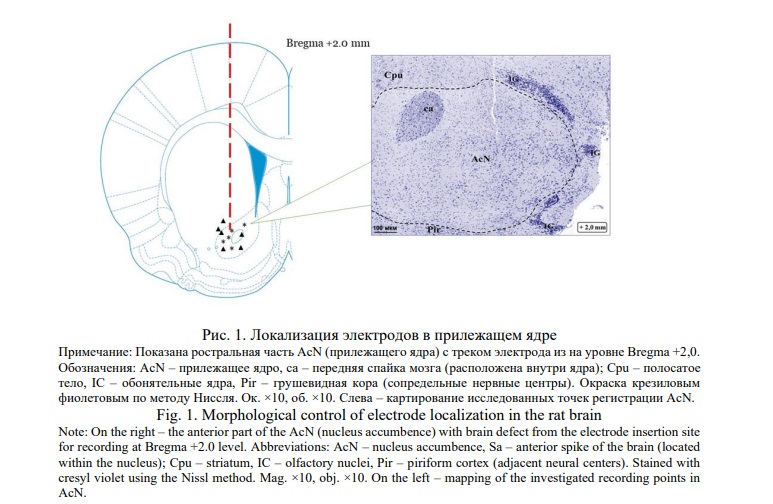
После проведения опытов по определению уровня дофамина мозг животных исследовали с целью установить локализацию кончика электрода в структуре мозга. Крыс после передозировки этаминалом натрия перфузировали, фиксировали в формалине, мозг выделяли и заливали целлоидином, делали фронтальные срезы и окрашивали крезиловым фиолетовым по методу Ниссля (рис. 1).
Применяя стереотаксический атлас мозга крыс [16], область локализации электродов верифицировали в гистологических срезах. Делали фронтальный разрез VTA на уровне «Bregma ‒5,3 мм», где структура хорошо выявляется. В этой области, паранигральном ядре, локализуются дофаминергические клетки. В прилежащем ядре делали фронтальный срез на уровне «Bregma +2,7 мм». Далее продолжали иссекать срезы на протяжении 0,7-1 мм до той области переднего мозга, в которой прилежащее ядро занимает наибольшую площадь (рис. 1). В этой части мозга передняя комиссура смещалась к верхней медиальной части ядра, тогда как в его наиболее крупной средней области располагался след электрода для регистрации (Рис. 1).
Для оценки соответствия распределений случайных величин гауссовым при статистической обработке данных использовали критерий нормальности Д'Агостино-Пирсона. Полученные данные в дальнейшем были представлены в виде (Q1, Me, Q3). Анализ данных проводили по критерию Краскела–Уоллиса с последующим множественным сравнением в тесте Данна. В работе использовали статистическую программу GraphPad Prizm 8.1 (GraphPad Software, США).
Результаты. У крыс была выработана реакция самостимуляции. При пороговых для самостимуляции значениях силы тока, от 50 до 80 мкА, наблюдали исследовательские реакции обнюхивания. Число нажатий педали варьировало от 65 до 130 нажатий за 5 мин опыта. При тестировании реакции самостимуляции использовали силу тока, на 50 выше пороговой, которая поддерживала длительную реакцию самостимуляцию. Целью настоящей работы было проанализировать особенности выброса дофамина, поэтому различия реакции самостимуляции в группах не оценивалось.
У крыс в свободном поведении проведено измерение внеклеточного уровня дофамина в NAc в ответ на раздражение области положительного подкрепления VTA. Было исследовано внеклеточное высвобождение дофамина у крыс CG и MD. При этом статистический анализ не выявил достоверных различий между группами CG и MD (рис. 2). В последующих исследованиях эти показания мы считали, как базовые, взятые за 100%, для изучения экстраклеточного дофамина в камерах Скиннера.
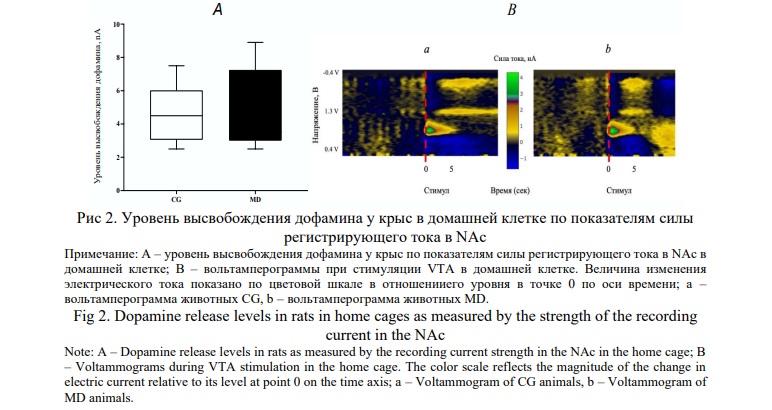
Эксперименты по определению изменений уровня дофамина в процессе реализации подкрепляющих свойств VTA не показало визуально отчетливых значений. В процессе реализации реакции самостимуляции интервалы между нажатиями педали различались от 0.2 до 9-10 с. В связи с этим в дальнейших исследованиях по определению изменений уровня дофамина использовали навязанную стимуляцию зоны самораздражения VTA пачкой импульсов с теми же параметрами в перерывах между сериями самостимуляции, отделяя при этом животное от педали прозрачной перегородкой.
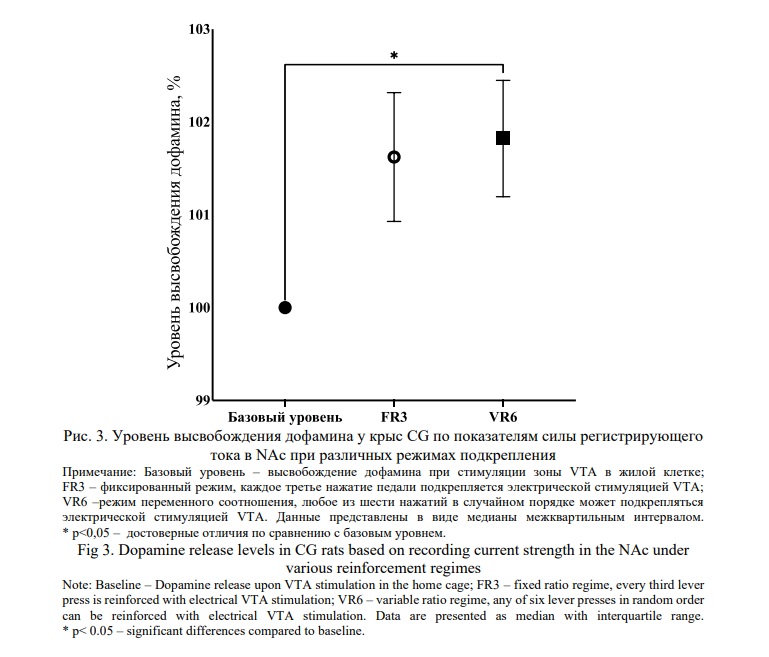
В эксперименте c определением уровня внеклеточного дофамина в NAc в ответ на стимуляцию зоны самораздражения VTA, определяемого по показателям электрохимического потенциала, критерий Краскела-Уоллиса выявил значимое влияние изучаемых режимов подкрепления FR3 и VR6 на уровень дофамина у крыс CG (Н(3) = 6. 804; р<0.05) по сравнению с базовым уровнем стимуляции. Множественное сравнение в тесте Данна выявило различия между медианами высвобождения уровня дофамина у животных с режимом подкрепления VR6 относительно базового уровня (n= 9, p<0.05), при сравнении режима подкрепления FR3 с базовым уровнем и режимом VR6 значимых отличий не выявлено (рис. 3).
В эксперименте c определением уровня внеклеточного дофамина в NAc в ответ на стимуляцию зоны самораздражения VTA, определяемого по показателям электрохимического потенциала, критерий Краскела-Уоллиса выявил значимые межгрупповые различия изучаемых режимов подкрепления на уровень дофамина у крыс MD (Н (3) = 15. 44; р<0.0001), а последующее множественное сравнение в тесте Данна выявило значительные различия между медианами высвобождения уровня дофамина у животных MD с режимом подкрепления VR6 относительно группы MD c базовым уровнем высвобождения дофамина (n=9, p<0.0001) и относительно группы с режимом подкрепления FR3 (n=9, p<0.01), что свидетельствует о формировании более выраженного аддиктивного поведения (рис. 4).
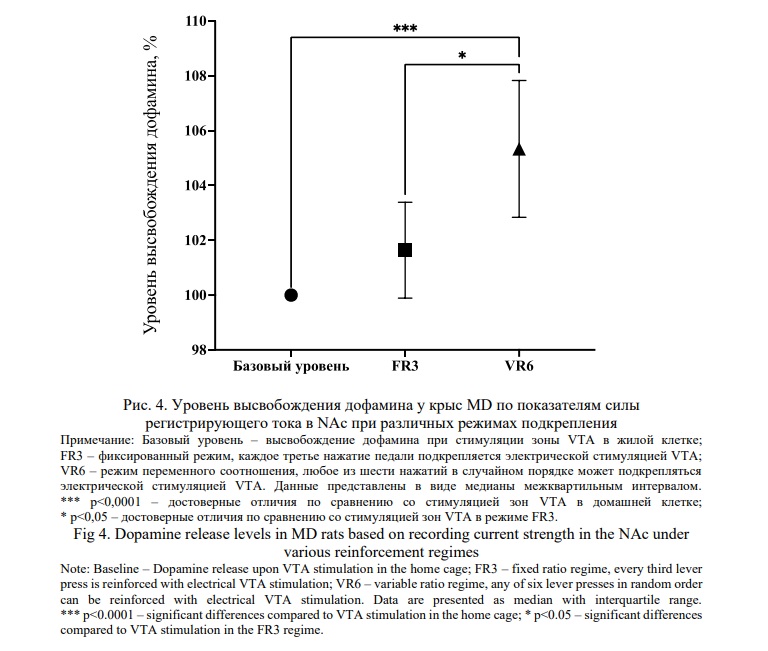
Запись кинетики высвобождения дофамина в NAc у контрольных и экспериментальных животных в ответ на электростимуляцию VTA показал, что величина подъёма уровня дофамина, определяемого по показателям электрохимического потенциала, у животных СG и MD при использовании режима подкрепления FR3 достоверно не отличалась. В то же время у животных MD наблюдался более выраженный дофаминовый ответ при использовании режима VR6 относительно базового уровня и режима FR3 (рис. 4, 5).
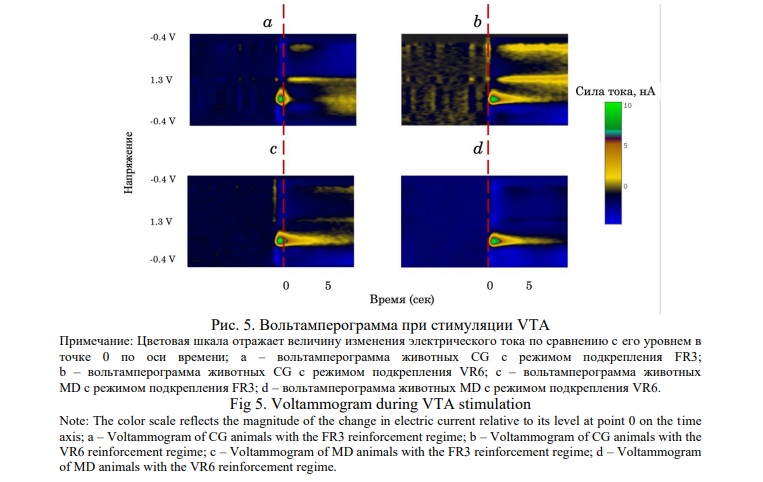
Обсуждение. В работе сделана попытка проанализировать особенности проявления аддиктивного состояния при моделировании элементов игровой зависимости у животных MD при самостимуляции VTA. Игровое расстройство, как было отмечено, имеют общие механизмы формирования с другими видами зависимости, наличие синдрома отмены, а также общие психофизиологические и нейробиологические характеристики [22, 23]. Ранее нами показано, что хронический стресс MD вызывает мотивацию крыс получить более значимое подкрепление в трехлучевом лабиринте при низкой его вероятности, когда возникал риск не получить вознаграждения, наблюдалось повышение импульсивности в поведении [2]. В настоящей работе проведено тестирование у животных реакции самостимуляции с использованием разных режимов подкрепления.
В работе показано, что стресс MD вызывает увеличение выброса дофамина, ассоциированной с режимом самостимуляции VR6, модели самостимуляции, приближенной к ситуации азарта и риска. Тестирование зоны VTA на наличие реакции самостимуляции позволяет судить о стимуляции подкрепляющей системы мозга мезолимбического дофаминергического нервного пути при регистрации внеклеточного дофамина в NAc. Аналогичные исследования по регистрации внеклеточного дофамина у MD не учитывали приуроченность зоны стимуляции к подкрепляющей системе мозга. Для проявления эффекта дофаминовой системы у животных MD в наших исследованиях дополнительно подавали условный сигнал аддиктивного состояния животного, применяя две камеры Скиннера с различными сенсорными характеристиками у CG и MD. Повышенная чувствительность дофаминовой системы, показанная в настоящей работе, связана с повышенной экспрессией тирозингидроксилазы и повышенной возбудимостью дофаминовых терминалей, что приводит к увеличению высвобождаемых везикулярных пулов и повышению чувствительности в ответ на стимулы [10]. Дофаминовые терминали проявляют повышенную возбудимость, возможно, из-за измененной экспрессии потенциал-зависимых кальциевых каналов и увеличения синтеза дофамина после длительного хронического стресса [24].
Известно, что активность дофаминовых нейронов повышается в ответ на условные сигналы, связанные с подкрепляющими и стрессовыми воздействиями [24]. У крыс MD в наших экспериментах наблюдалось повышение чувствительности дофамина на условный сигнал аддиктивного поведения при моделировании состояния азарта и риска, элементов игровой зависимости. Гиперчувствительность системы дофамина на аддиктивные стимулы среды потенциально могут привести к сдвигу величины воспринимаемого вознаграждения и активировать животное в ответ на восприятие большего по значимости подкрепления [25]. В настоящем исследовании мы определяли внеклеточный уровень дофамина в NAc, который, как известно, участвует в механизмах стресс-реактивности, тревожности, депрессии и мотивации, а также процессах, связанных с подкреплением [26]
По-видимому, в онтогенезе у грызунов существуют особые критические периоды чувствительности структур головного мозга к формированию отдаленных проявлений аддиктивного поведения, которые затрагивают исследованные в работе дни MD [7]. Важные биологические стрессоры в онтогенезе могут привести к аналогичным нейрохимическим и поведенческим изменениям [27].
В работе показано, что стресс материнской депривации в раннем онтогенезе вызывает увеличение выброса внеклеточного дофамина в прилежащем ядре у половозрелых крыс при использовании режима переменного соотношения самостимуляции, приближенной к ситуации азарта и риска. При этом в работе стимулировали зону награды VTA, где локализуются дофаминергические клетки и стимулируют выброс дофамина в прилежащем ядре. Аналогичные исследования по регистрации внеклеточного дофамина у MD не учитывали приуроченность зоны стимуляции к подкрепляющей системе мозга. Проведенные эксперименты позволяют анализировать дофаминергические механизмы состояния азарта и риска в реальном времени. В то же время имеется ограничение используемого метода, которое состоит в необходимости проведения операций по вживлению электродов в мозг. Комплексная оценка поведенческих, биохимических и электорофизиологических проявлений аддиктивного поведения позволит обоснованно проводить поиск новых мишеней для фармакологического воздействия при моделировании элементов игровой зависимости. Этими мишенями, по-видимому, являются рецепторы следовых аминов, дофаминовые, серотониновые, грелиновые, кортиколибериновые, кисспептиновые рецепторы эмоциогенных структур мозга, которые запускают эмоционально окрашенную реакцию в ситуации, приближенной к риску и азарту у человека, поддерживают ее, способствуя формированию или же возобновлению провоцирующих игровых факторов (риска и азарта). Перспективы коррекции указанных состояний видятся в блокаде указанных подтипов рецепторов с помощью фармакологических соединений традиционной химической структуры и генноинженерных продуктов на основе плазмидных ДНК, позволяющих направленно синтезировать специфические белки-антагонисты рецепторов.
Заключение. Стресс материнской депривации в раннем онтогенезе вызывает повышение высвобождения дофамина в прилежащем ядре в ответ на стимуляцию зоны положительного подкрепления, которая сопровождается подачей условного сигнала аддиктивного состояния в модели самостимуляции при использовании режима переменного соотношения. Материнская депривация вызывает повышение выброса дофамина при моделировании состояния риска и азарта, что создает перспективы для изучения механизмов дизонтогенеза в уязвимости к действию подкрепляющих стимулов, вызывающих игровое расстройство поведения (например, офлайн- и онлайн-игры, в которые можно играть на любом цифровом устройстве: мобильном телефоне, компьютере).
Информация о финансировании
Работа выполнена в рамках государственного задания FGWG-2025-0020 «Поиск молекулярных мишеней для фармакологического воздействия при аддиктивных и нейроэндокринных нарушениях с целью создания новых фармакологически активных веществ, действующих на рецепторы ЦНС».














Список литературы