
Роль генов глутатион-зависимых ферментов антиоксидантной системы в выживаемости и достижении долголетия
Aннотация
Актуальность: Долголетие человека характеризуется способностью поддерживать внутриклеточный гомеостаз в изменяющихся с возрастом условиях. Эффективность протекания многих редокс-зависимых эндогенных процессов зависит от структурно-функциональных особенностей генов, связанных с глутатионом. Цель исследования:Исследование роли полиморфных локусов генов глутатион-зависимых ферментов антиоксидантной системыGSR (rs1002149), GSTP1 (rs1695), GPX1 (rs1050450), GPX4 (rs713041) и GPX8 (rs381852) в выживаемости и достижении долголетия при физиологическом и патологическом старении. Материалы и методы:Выборка из 3218 человек, русских, башкир и татар по этнической принадлежности, включала лиц среднего возраста (18-59 лет), старшего возраста (60-89 лет) и долгожителей (90-114 лет). Информация о статусе выживаемости и окончательном диагнозе получена для 1656 лиц в возрасте старше 45 лет. Полиморфные генетические маркеры проанализированы с использованием TaqMan-ПЦР технологии. Результаты:Установлены генетические маркеры долголетия и выживаемости, включающие аллель GPX4 (rs713041) Т (мужчины, OR=1,66, P=0,006; HR=0,81, Р=0,02), генотип GPX4 (rs713041) СТ (общая группа, OR=1,33, P=0,004; коморбидный статус, HR=0,6, P=0,02), а также сочетание аллелей GPX1 (rs1050450) G и GSTP1 (rs1695) A (женщины, русские по этнической принадлежности, OR=4,15, PFDR=0,04; общая группа, HR=0,5, Р=0,02). В этнической группе русских с долголетием среди женщин ассоциирован аллель GPX8 (rs381852) A (OR=2,52, P=0,005); в гомозиготном и гетерозиготном варианте он встречался в девяти сочетаниях, способствующих долголетию (в комбинации с аллелями GPX1 (rs1050450) G и GSTP1 (rs1695) A показаны наиболее значимые статистические показатели – OR=4,07, PFDR=0,006). Сочетание GPX1 (rs1050450) GG,GPX4 (rs713041) C и GSR (rs1002149) T ассоциировано с долголетием башкир (OR=7,98, PFDR=0,02). Генотип GSR (rs1002149) ТТ является маркером смертности от всех причин (HR=1,47, Р=0,02). Со смертностью от цереброваскулярных заболеваний в преклонном возрасте связаны генотипGPX1 (rs1050450) AG (HR=1,42, Р=0,003) и сочетание аллелейGPX1 (rs1050450) G и GSTP1 (rs1695) A (HR=1,4, Р=0,02). Заключение:В генах глутатион-зависимых ферментов антиоксидантной системывыявлены общие и этноспецифические генетические предикторы выживаемости в преклонном возрасте, смертности с учетом клинических данных и долголетия
Ключевые слова: долголетие, старение, возраст-зависимые заболевания, глутатион-зависимые ферменты антиоксидантной системы, генетический полиморфизм, ген-генные взаимодействия, выживаемость
Введение. Долголетие представляет собой уникальный феномен исключительной продолжительности жизни, значительно превышающей средний популяционный показатель. Важным условием успешного онтогенеза является высокая степень адаптации организма к стрессогенным факторам среды и сохранение постоянства внутренней среды организма [1]. Окислительно-восстановительные процессы выступают в качестве основных регуляторов внутриклеточного гомеостаза [2]. Ключевую роль в редокс-процессах играет глутатион (GSH) – трипептид, состоящий из цистеина, глицина и глутаминовой кислоты, который в высоких концентрациях обнаруживается практически во всех клетках организма [3]. Глутатион является мобильным донором свободных электронов и присутствует в клетке в двух формах – восстановленной (GSH) и окисленной (GSSG). Именно соотношение GSH/GSSG определяет окислительно-восстановительный статус клетки. Высокая концентрация GSH способствует эффективному восстановлению аминокислотных остатков, при этом сам глутатион окисляется до GSSG. Взаимодействуя с кислородными радикалами, связывая ксенобиотики, стабилизируя клеточные мембраны, GSH является универсальным антиоксидантом [4].
Система метаболизма глутатиона как одна из важнейших регуляторных механизмов, обеспечивающих внутриклеточный гомеостаз и клеточное выживание, рассматривается в рамках глутатионовой теории старения [5, 6]. Разнообразные внешние и поведенческие факторы, такие, как несбалансированная диета, воздействие ксенобиотиков, могут приводить к истощению глутатиона и дисбалансу его окисленной и восстановленной форм [7]. Более того, с возрастом снижается способность организма синтезировать глутатион [7]. Большинство возрастных хронических дегенеративных заболеваний связаны с низким уровнем глутатиона и ферментов, участвующих в его метаболизме [6, 8].
Ферментный глутатионовый комплекс антиоксидантной системы включает глутатионредуктазу (GSR), семейство глутатионпероксидаз (GPx) и глутатионтрансферазу (GST), представленную несколькими изоформами. Данные компоненты метаболизма глутатиона являются сенсорами и регуляторами внутриклеточного редокс-статуса [8-13]. Глутатионредуктаза (Glutathione S-Reductase, GSR) – перманентно активированный флавопротеин, восстанавливающий окисленный дисульфид глутатиона (GSSG) до сульфгидрильной формы GSH. Таким образом, GSR поддерживает концентрацию восстановленного глутатиона, и его активность повышается в условиях окислительного стресса [9]. Различные изоформы GPx восстанавливают перекиси водорода (Н2О2), используя глутатион в качестве косубстрата. Тем самым GPx регулируют концентрацию данного промежуточного кислородного метаболита, который является не только окислителем, но и важной сигнальной молекулой, участвующей во множестве клеточных процессов. Глутатионпероксидаза 1 (GPx1) является самой распространенной и наиболее активной в отношении восстановления гидроперекисей изоформой [10]. Глутатионпероксидаза 4 (GPx4) восстанавливает гидроперекиси сложных липидов, участвуя таким образом в поддержании целостности мембран и ферроптозе [11]. Глутатионпероксидаза 8 (GPx8) – трансмембранный белок эндоплазматического ретикулума, участвующий в окислительном фолдинге белков и регулировании концентрации ионов Са+ [12]. Глутатион-S-трансферазы (GST) – крупнейшее суперсемейство ферментов второй фазы детоксикации. Данные белки обеспечивают связывание глутатиона с различными токсичными соединениями, нейтрализуя их электрофильные свойства и способствуя удалению из клетки [8]. Кроме того, S-глутатионилирование опосредует редокс-регуляцию активности каталитических белков, в частности, ингибируя, или, наоборот, активизируя связывание транскрипционных факторов с ДНК [13].
Изменение активности фермента и соответствующий эффект в отношении устойчивости или уязвимости к эндогенным и экзогенным факторам могут быть связаны с генетическим полиморфизмом в кодирующем гене. Так, для полиморфного локуса rs1002149 гена GSR была установлена ассоциация с уровнем активности фермента GSR и с выживаемостью среди супердолгожителей [14]. Полиморфизм rs1050450, приводящий к нарушению ферментативной активности гена GPX1, ассоциирован с развитием возраст-зависимых патологий сердечно-сосудистой системы, онкологических заболеваний [10]. Полиморфизм rs713041, расположенный в 3’-регуляторной области гена GPX4, модулирует степень связывания фермента с селеном и, соответственно, его экспрессию, и ассоциирован с риском развития заболеваний возраста [15]. Полиморфный вариант rs381852 гена GPX8 ассоциирован на уровне GWAS с раком яичников [16]. Показана ассоциация полиморфного варианта rs1695 в гене GSTP1 с некоторыми видами рака, болезнями сердечно-сосудистой системы, органов дыхания [17, 18]. Таким образом, функциональные полиморфные варианты генов глутатионового обмена ассоциированы с болезнями возраста и могут быть вовлечены в формирование молекулярно-генетического фона для физиологической адаптации организма, способствующей выживаемости.
Цель исследования заключалась в изучении роли полиморфных локусов генов глутатион-зависимых ферментов антиоксидантной системы GSR (rs1002149), GSTP1 (rs1695), GPX1 (rs1050450), GPX4 (rs713041) и GPX8 (rs381852) в выживаемости и достижении долголетия среди населения республики Башкортостан с учетом пола, этнической принадлежности и клинико-патологического диагноза.
Материалы и методы исследования. Исследование одобрено Локальным этическим комитетом ИБГ УФИЦ РАН (Протокол № 1 от 4 ноября 2007 г.) и выполнено в соответствии с этическими принципами проведения медико-биологических исследований с участием человека в качестве субъекта, закрепленными в Хельсинской декларации Всемирной медицинской ассоциации (World Medical Association, 2013).
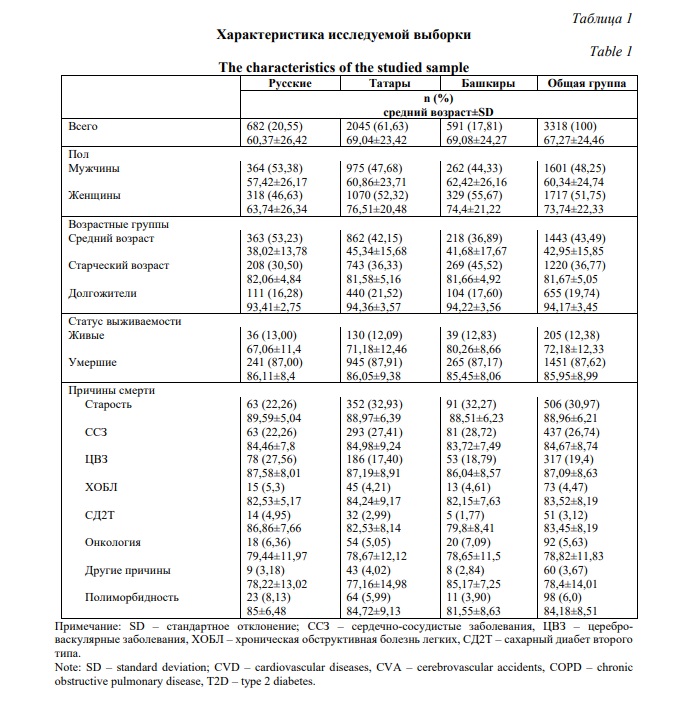
Общая выборка включала 3218 не родственных между собой мужчин и женщин в возрасте от 18 до 114 лет, русских, башкир и татар по этнической принадлежности, проживающих в Республике Башкортостан. После получения информированного добровольного согласия на участие в исследовании проводили анкетирование, включающее также данные анамнеза. Для лиц среднего возраста (18-59 лет) на момент забора биологического материала исключались инфаркт миокарда, инсульт, аутоиммунные расстройства, сахарный диабет и онкологические заболевания. Для участников старшего возраста (60-89 лет) допускалось наличие в анамнезе атеросклероза. В группу долгожителей включали всех лиц старше 90 лет.
Получена информация о статусе выживаемости и окончательном диагнозе 1656 исследуемых лиц старше 45 лет. На основании верифицированных данных о доживаемости, полученных к концу 2024 года, вся выборка дифференцирована по возрасту (группы лиц среднего, старшего возраста и долгожителей) и по причинам смертности от различных патологий (на основании данных заключений о смерти). Общая выборка и выделенные группы сравнения охарактеризованы в таблице 1.
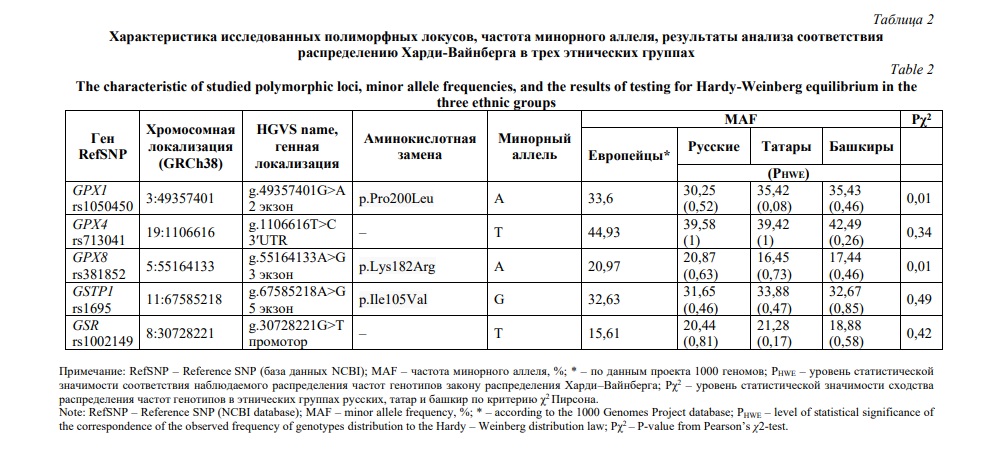
Отобранные для исследования гены кодируют ключевые ферменты антиоксидантной системы, использующие глутатион в качестве косубстрата [13]. Полиморфные маркеры генов GSR (rs1002149), GSTP1 (rs1695), GPX1 (rs1050450), GPX4 (rs713041) и GPX8 (rs381852) были подобраны с использованием компьютерных баз данных GWAS Catalog (https://www.ebi.ac.uk/gwas/, дата обращения сентябрь 2023 г.), NCBI (The National Center for Biotechnology Information, https://www.ncbi.nlm.nih.gov/, дата обращения сентябрь 2023 г.), Ensembl genome browser (https://www.ensembl.org/index.html, дата обращения сентябрь 2023 г.). Функциональная значимость отобранных для исследования SNP анализировалась в программах RegulomeDB (https://regulomedb.org, дата обращения сентябрь 2023 г.) и Open Targets Genetics (https://genetics.opentargets.org/, дата обращения сентябрь 2023 г.). Данные полиморфные варианты локализованы в кодирующих или регулирующих сайтах генов и связаны с изменением генной экспрессии, способны оказывать специфические эффекты в отношении сигнальных путей, в которые вовлечены белковые продукты изучаемых генов [8, 11, 12, 19, 20], а также являются генетическими маркерами ряда возраст-ассоциированных заболеваний [9, 13, 16, 18, 19, 21-27]. Характеристика исследованных SNP представлена в таблице 2.
Образцы ДНК получены путем фенольно-хлороформной экстракции из 8 мл цельной венозной крови. Генотипирование проведено методом ПЦР в реальном времени с использованием наборов праймеров и целевых флуоресцентных TaqMan-зондов («ДНК-Синтез», Россия). Распределение частот генотипов было проверено на соответствие равновесию Харди-Вайнберга в контрольной группе. Различие в распределении частот генотипов между этническими группами для каждого SNP оценивалось с помощью критерия χ2 Пирсона (SPSS Statistics v. 22.0.). Ассоциации исследуемых SNP с возрастом анализировались методом логистической регрессии с построением доминантной, кодоминантной, сверхдоминантной, рецессивной и лог-аддитивной генетических моделей (программная среда R v. 4.4.1., пакет SNPassoc v. 2.1-2, https://cran.r-project.org/, [28]). Наиболее подходящая генетическая модель выбиралась для каждого SNP, статистически значимо связанного с исследуемым признаком, на основе наименьшего значения информационного критерия Акаике (AIC). Ассоциированные со старением и долголетием комбинации аллелей и генотипов изучаемых генов устанавливались с использованием программы APSampler (v.3.6.0, http://apsampler.sourceforge.net, [29]). Корреляция отдельных полиморфных локусов и их сочетаний с общей выживаемостью и при различных клинических фенотипах устанавливалась с использованием регрессии пропорциональных рисков Кокса; кривые выживаемости визуализировались с использованием графиков Каплана–Майера (SPSS Statistics v. 22.0.). Все расчеты проводились с поправкой на пол и этническую принадлежность. Детальное описание методов и подходов статистического анализа описаны нами ранее [30].
Результаты и их обсуждение. В этнических группах русских, татар и башкир, которые являются наиболее распространенными в структуре населения Республики Башкортостан, охарактеризованы полиморфные локусы генов ферментов антиоксидантной системы, связанных с глутатионом. Наблюдаемое распределение частот генотипов в каждой исследованной этнической группе соответствовало теоретически ожидаемому равновесному распределению Харди-Вайнберга (PHWE>0,05, Табл. 2). Сравнительный анализ распределения частот генотипов по каждому исследованному SNP выявил различия между этническими группами по полиморфным локусам GPX1 (rs1050450) и GPX8 (rs381852) (Р=0,01, табл. 2).
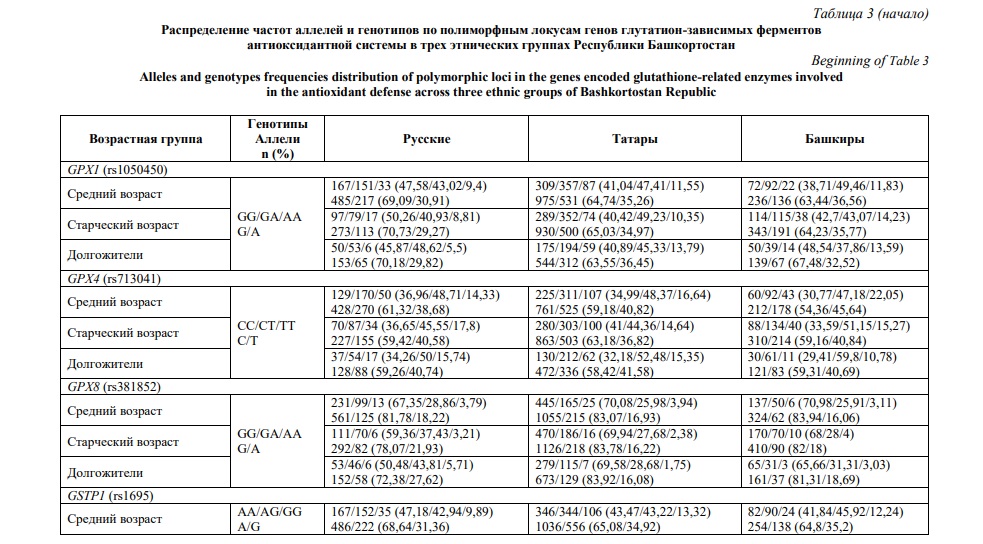
Распределение частот аллелей и генотипов в этнических группах с учетом возрастной градации приведено в таблице 3. Ассоциации между возрастными группами и полиморфными локусами оценивались с помощью регрессионного анализа и представлены в рамках различных генетических моделей в таблице 4. Наибольшее число ассоциаций продемонстрировано для полиморфного локуса rs713041 гена GPX4. В общей выборке частота генотипа GPX4 (rs713041) СТ оказалась выше в группе долгожителей, относительно таковой среди лиц среднего (P=0,021) и преклонного (P=0,004) возраста. Ассоциация аллеля GPX4 (rs713041) Т с долголетием, относительно группы лиц преклонного возраста, выявлена в общей группе (P=0,007) и в группе татар (P=0,004). Более того, в группе мужчин данный аллель, по которому также показана ассоциация с долголетием (P=0,006), был установлен как предиктор общей выживаемости (HR=0,81, Р=0,02, Рис. 1А). В то же время у башкир снижена частота генотипа GPX4 (rs713041) ТТ среди лиц преклонного возраста (P=0,04), долгожителей (P=0,008), и особенно женщин старше 90 лет (P=0,001). Генотип GPX8 (rs381852) GA ассоциирован с долголетием в общей группе (P=0,03). В группе женщин русской этнической принадлежности с долголетием ассоциированы генотипы GPX1 (rs1050450) GA (P=0,02) и GSTP1 (rs1695) AG (P=0,008), а также аллель GPX8 (rs381852) A (P=0,005). Частота генотипа GSR (rs1002149) ТТ снижена среди татар долгожителей (P=0,03); данный генотип также был идентифицирован как маркер повышенного риска смертности от всех причин (HR=1,47, Р=0,02, рис. 1Б).
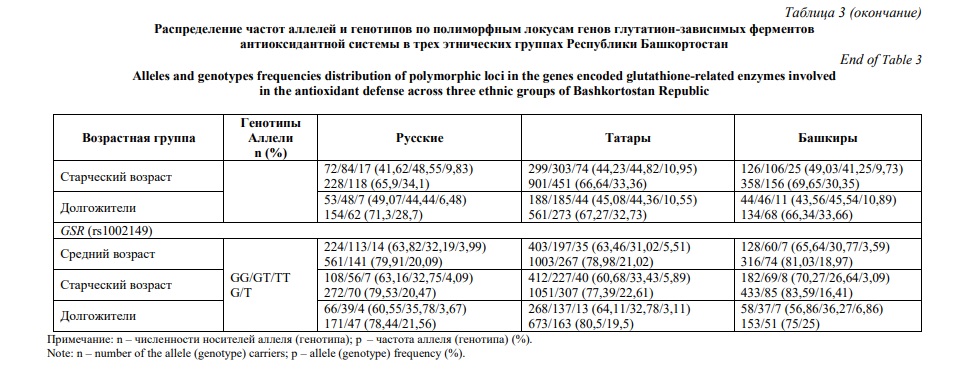
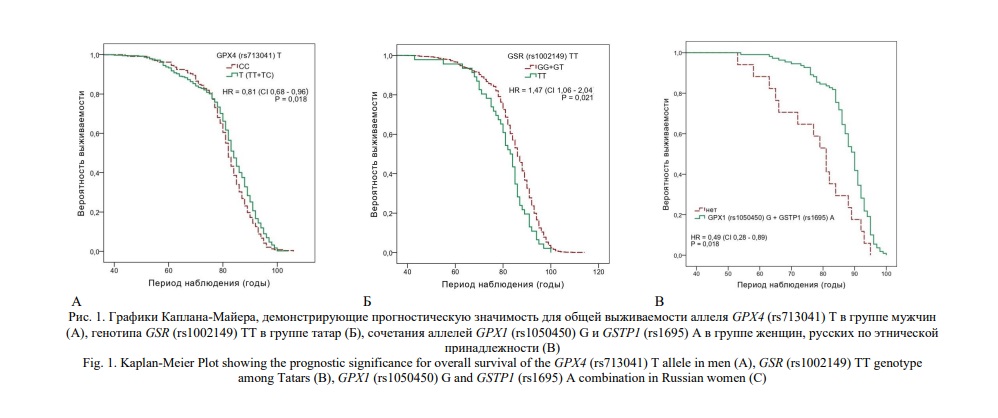
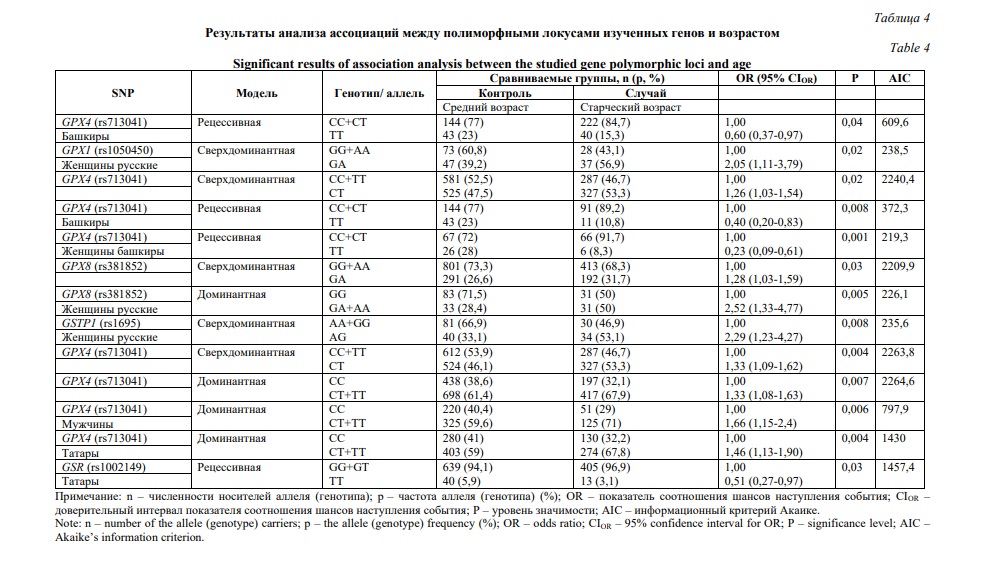
Выполнен поиск информативных ген-генных сочетаний, связанных с достижением преклонного возраста и долголетия. Установлено 11 комбинаций аллелей и генотипов, ассоциированных с долголетием в группе женщин (OR>3, PFDR<0,05) (табл. 5). Среди башкир, достигших долголетия, вероятность встречаемости сочетания GPX1 (rs1050450) GG +GPX4 (rs713041) C + GSR (rs1002149) T в восемь раз выше, чем в группе лиц среднего возраста (PFDR=0,02). В группе русских аллели GPX1 (rs1050450) G, GPX8 (rs381852) A и GSTP1 (rs1695) A формируют 10 различных комбинаций, с наиболее значимым статистическим показателем для сочетания GPX1 (rs1050450) G + GPX8 (rs381852) A + GSTP1 (rs1695) A (PFDR=0,006). Сочетание аллелей GPX1 (rs1050450) G и GSTP1 (rs1695) A, ассоциированное с долголетием (PFDR=0,05), оказалось значимым для общей выживаемости (HR=0,5, Р=0,02, Рис. 1В).
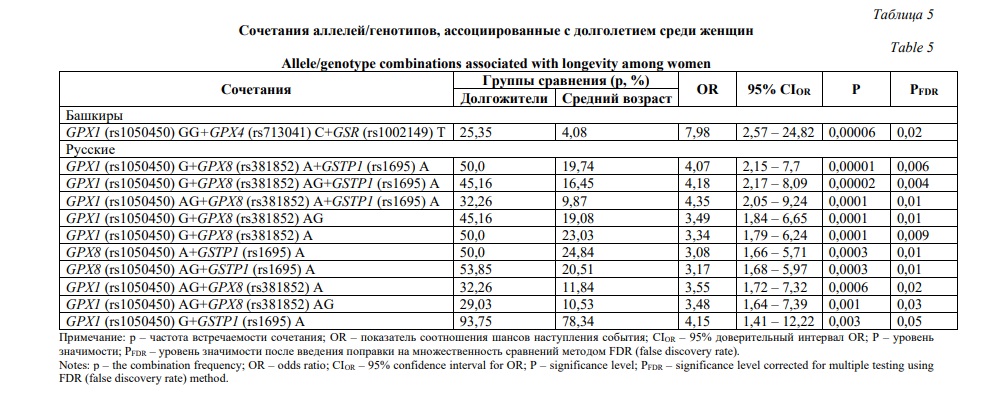
Установленные в результате построения регрессионных моделей и ген-генных комбинаций генетические маркеры долголетия были выбраны в качестве предикторов для проведения анализа выживаемости в группах, сформированных на основе сведений о причинах смерти исследуемых лиц. Маркерами риска смертности при цереброваскулярных заболеваниях оказались генотип GPX1 (rs1050450) AG (HR=1,42, Р=0,003, Рис. 2А) и сочетание аллелей GPX1 (rs1050450) G и GSTP1 (rs1695) A (HR=1,4, Р=0,02, Рис. 2Б). С выживаемостью при коморбидном состоянии ассоциирован генотип GPX4 (rs713041) СТ (HR=0,6, P=0,02, Рис. 2В).
Таким образом, на обширной выборке, включающей мужчин и женщин трех крупных этнических групп русских, татар и башкир, проживающих на территории Республики Башкортостан, изучено возраст-зависимое изменение спектра частот аллелей и генотипов по полиморфным локусам в генах глутатион-связанных ферментов антиоксидантной системы GSR (rs1002149), GSTP1 (rs1695), GPX1 (rs1050450), GPX4 (rs713041) и GPX8 (rs381852). Исследованная выборка сформирована из коренных жителей Республики Башкортостан и имеющих тем самым сходные тенденции в отношении адаптации к экологическим и климатогеографическим условиям. В то же время были выявлены этнические особенности в распространении частот генотипов по полиморфным локусам генов GPX1 (rs1050450) и GPX8 (rs381852) (табл. 2). Кроме того, установлены как общие, так и этноспецифические ассоциации разных генетических маркеров с долголетием и выживаемостью в преклонном возрасте (табл. 4-5, рис. 1-2).
Аллель (rs1050450) А ассоциирован со сниженной активностью гена GPX1 и соответственно с низким уровнем белка, с понижением концентрации селена в крови, а также с рядом патологий возраста в разных популяциях мира [19]. Полученная в группах женщин русской и башкирской этнической принадлежности ассоциация аллеля GPX1 (rs1050450) G с долголетием подтверждает участие данного полиморфного маркера в формировании фенотипа долгожителя, устойчивого к возрастзависимым заболеваниям. Гетерозиготный генотип GPX1 (rs1050450) АG оказался вовлечен в формирование патологического фенотипа, связанного с цереброваскулярными событиями и смертностью (рис. 2А). Интересно, что наличие даже одного аллеля GPX1 (rs1050450) А повышало риск и степень развития заболеваний возраста [19]. При этом на модельных объектах показано участие нормальной и высокой активности GPх1 в защите от окислительного стресса именно в тканях нервной системы [12]. Можно предположить, что полиморфный маркер GPX1 (rs1050450) вовлечен в формирование адаптивных механизмов при развитии сосудистых патологий головного мозга на поздних этапах онтогенеза, способствующих выживаемости в очень преклонном возрасте.
Ассоциация полиморфизма GPX1 (rs1050450) с долголетием и показателями смертности дополнительно наблюдалась и в сочетании с GSTP1 (rs1695) (табл. 5, рис. 1В, 2Б), что может свидетельствовать о критической роли GPХ1 и GSTР1 в качестве регуляторов редокс-гомеостаза для клеточной выживаемости на поздних этапах онтогенеза. Полиморфизм rs1695 гена GSTP1 ассоциирован с сердечно-сосудистыми заболеваниями и раком [13, 18, 21, 22], а также с болезнью Альцгеймера [23]. В то же время было установлено снижение частоты аллеля GSTP1 (rs1695) G с возрастом у итальянских долгожителей [31], что согласуется с нашими результатами, согласно которым аллель GSTP1 (rs1695) А в сочетании с другими аллелями генов семейства GPx ассоциирован с долголетием (табл. 5). Таким образом, GSTР1 ассоциирован с физиологическими и патологическими возрастными процессами. Это обусловлено тем, что GST способен взаимодействовать с различными киназами и регулировать важнейшие внутриклеточные сигнальные каскады – JNK, ASK1, MAP, участвуя тем самым в апоптозе и стресс-реакциях [8].
GPx4 также является важнейшим регулятором выживаемости клетки, так как способна напрямую восстанавливать гидроперекиси липидов и холестерина [11], запускать AIF-опосредованную клеточную гибель [32] и ингибировать ферроптоз [11, 33]. Из всех известных изоформ GPx только для GPx4 была установлена связь дефицита данного фермента с выживаемостью эмбрионов лабораторных мышей [34]. Полиморфный локус GPX4 (rs713041) связан с содержанием селенопротеинов в клетке – в условиях дефицита селена концентрация GPx4 лучше поддерживалась у носителей генотипа СС; данный полиморфизм тем самым был определен в качестве маркера восприимчивости к болезням [24]. Так, аллель GPX4 (rs713041) T ассоциирован с риском развития инсульта, гипертонии, рака [15]. В нашей работе для полиморфного локуса GPX4 (rs713041) получено наибольшее число ассоциаций, в то же время для разных этнических групп маркеры долголетия различались. Аллель GPX4 (rs713041) С, для которого показана защитная роль в отношении окислительного повреждения ДНК при нормальном уровне селена [20], ассоциирован с долголетием у башкир, как индивидуальный генетический маркер, так и в мультилокусных паттернах (табл. 4, 5). Аллель GPX4 (rs713041) Т оказался ассоциирован с долголетием и общей выживаемостью мужчин татарской этнической принадлежности (табл. 4, рис. 1А). Принимая во внимание роль данного полиморфизма в обмене и функционировании селенсодержащих ферментов, можно предположить, что исторически сложившиеся особенности рациона и экологических условий проживания могли сформировать этноспецифические адаптационные молекулярные механизмы [35].
Результаты исследований GPх8 демонстрируют его участие в различных физиологических процессах, таких как антиоксидантная защита, стресс эндоплазматического ретикулума, апоптоз, регуляция гомеостаза кальция, внутриклеточный сигналинг, а также в опухолеобразовании [36]. В то же время отсутствуют работы по изучению полиморфных вариантов гена GPX8. Методом GWAS была установлена ассоциация rs381852 с раком яичников [16]. В нашем исследовании аллель GPX8 (rs381852) А ассоциирован с долголетием женщин, русских по этнической принадлежности, как отдельно, так и в сочетаниях (табл. 4, 5).
Функциональный полиморфный вариант GSR (rs1002149) Т ассоциирован с высоким уровнем GSH, который в свою очередь коррелирует с благоприятным физиологическим фоном, способствующим долголетию и устойчивости к развитию заболеваний [9, 14]. Более того, низкая концентрация сывороточного GSH является предиктором когнитивных нарушений, болезни Альцгеймера, мультиморбидного статуса и старения в целом [10, 25]. С другой стороны, аллель (rs1002149) T является аллелем риска для рака груди [9], хронической обструктивной болезни легких [26], коррелирует со снижением минеральной плотности костной ткани в постменопаузе [27] и более того, с выживаемостью среди супердолгожителей [14]. Таким образом, полученные для этнической группы татар результаты, демонстрирующие снижение частоты встречаемости генотипа (rs1002149) ТТ среди долгожителей (табл. 4) и повышенный риск смертности (Рис. 1Б), согласуются с таковыми в других европейских популяциях. В то же время у башкир способствующее долголетию сочетание включает аллель (rs1002149) T (табл. 5); каждый элемент сочетания таким образом ассоциирован с высокой активностью кодируемого им фермента, и тем самым вовлечен в поддержание высокого уровня восстановительного потенциала глутатионового компонента антиоксидантной системы.
Заключение. В данном исследовании мы оценили вклад полиморфных вариантов генов глутатион-зависимых ферментов антиоксидантной системы GSR (rs1002149), GSTP1 (rs1695), GPX1 (rs1050450), GPX4 (rs713041) и GPX8 (rs381852) в формирование фенотипа долгожителя среди основных этнических групп населения Республики Башкортостан. Для аллеля GPX4 (rs713041) С, связанного с лучшей эффективностью работы GPx4, установлено наибольшее число ассоциаций с долголетием и выживаемостью в этнической группе башкир. В то же время в группе татар протективным в отношении выживаемости оказался аллель GPX4 (rs713041) Т, ассоциированный с риском развития многофакторных патологий; этот результат требует дальнейшего изучения этноспецифических молекулярных и биохимических процессов. В группе женщин, русских по этнической принадлежности, аллель GPX8 (rs381852) A ассоциирован с долголетием как индивидуально, так и в сочетании с другими генетическими маркерами. В этнической группе татар аллель GSR (rs1002149) Т, который является маркером риска многих возраст-зависимых заболеваний, в гомозиготном варианте повышает риск смертности от всех причин. Среди женщин, русских по этнической принадлежности, аллели GPX1 (rs1050450) G и GSTP1 (rs1695) A, связанные с нормальной и высокой активностью кодируемых белковых продуктов, способствуют достижению долголетия, но в то же время демонстрируют эффект антагонистической плейотропии, поскольку ассоциированы с риском смертности именно в возрасте долголетия, и особенно при развитии цереброваскулярных патологий.
Таким образом, пол и этническая принадлежность способны модифицировать эффекты работы генов глутатион-зависимых ферментов клеточной антиоксидантной защиты. Кроме того, изученные полиморфные локусы выступают маркерами уязвимости или устойчивости к развивающимся с возрастом патологиям, и могут зависеть от гормонального и физиологического статуса, характерного для определенного возрастного этапа. При этом, поскольку эффекты отдельных генов реализуются в многоуровневых и разветвленных сигнальных молекулярных путях, их влияние на выживаемость может меняться в зависимости от генетического окружения.
В целом, исследованные полиморфные локусы ассоциированы с выживаемостью при различных физиологических и патологических состояниях, а также с долголетием, и являются частью сложной динамичной сети молекулярных внутриклеточных взаимодействий.
Информация о финансировании
Исследование выполнено за счет гранта Российского научного фонда № 24-25-00179 (https://rscf.ru/project/24-25-00179/).














Список литературы