
Вторичные метаболиты Reynoutria sachalinensis – потенциальный резерв фармакологических препаратов (обзор)
Aннотация
Актуальность: Литературный обзор посвящен многолетнему травянистому растению Reynoutria sachalinensis семейства Polygonaceae. Во многих странах данный вид считается инвазивным и ведется поиск не только методов борьбы с ним, но и вариантов его использования, как лекарственного, медоносного, декоративного, пищевого и кормового растения. Цель исследования:Систематизировать современные научные данные в области химического состава R. sachalinensis, механизмов действия его основных биологически активных соединений и обоснованию их терапевтического использования. Материалы и методы:Для достижения поставленной цели проводился анализ источников отечественной и иностранной литературы по данной проблеме за период с 2000 г. по 2025 г. Использовались базы данных научных электронных библиотек Google Scholar, eLibrary, PubMed, Web of Science, ScienceDirect, Scopus. Результаты:В обзоре обобщены результаты исследований в области изучения химического состава, биологической активности как различных экстрактов, так и индивидуальных соединений, а также механизмов действия основных биологически активных веществ R. sachalinensis. В настоящее время из данного вида выделено и идентифицировано более 277 соединений. Метаболиты R. sachalinensis относятся к фенольным соединениям, тритерпеноидам, азотсодержащим соединениям, пигментам и алифатическим спиртам. В исследованиях последних десятилетий показано, что суммарные экстракты и высокоочищенные фракции соединений из различных органов R. sachalinensis обладают антиоксидантной, противовирусной, фунгицидной и противоопухолевой активностью, ингибирующим действием на β-глюкозидазу, ацетилхолинэстеразу, ксантиноксидазу и нейропротективными свойствами. Знание пространственной структуры комплексов клеточных белков, мембранных рецепторов и растительных вторичных метаболитов, их лигандов раскрывает механизм действия биологически активных веществ, но не исключает необходимости его подтверждения и проведения эксперимента in vitro иin vivo. Заключение:Растительное сырье R. sachalinensis можно рассматривать как источник биологически активных метаболитов, имеющих широкий спектр использования
Ключевые слова: рейнутрия сахалинская, вторичные метаболиты, цитотоксичность, противовирусное действие, молекулярный докинг
Введение. Лекарственные растения играют важную роль в мировом здравоохранении и глобальной экономике [1]. В настоящее время индустрия фитотерапии переживает активный рост [2]. Только в США более 25% рецептурных лекарств содержат активные вещества, полученные из растений [3]. В России доля рынка фитопрепаратов меньше по сравнению со странами Евросоюза [4, 5, 6]. При этом Российская Федерация обладает определенным потенциалом для производства фитопрепаратов [7, 8]. Исследования, проводимые в области поиска и изучения механизма действия вторичных метаболитов лекарственных растений, являются актуальными [9].
Reynoutria sachalinensis (F. Schmidt) Nakai (син. Polygonumsachalinensis F. Schmidt ex Maxim., Fallopiasachalinensis (F. Schmidt) Ronse Decr.) – Рейнутрия сахалинская – многолетнее травянистое растение высотой до 4 метров с крупными овально-продолговатыми, с сердцевидным основанием и острой или заостренной верхушкой листьями, длина которых варьирует от 15 до 45 см. [10, 11, 12]. Данный вид принадлежит семейству Polygonaceae [13]. Корневище утолщенное, ветвистое. Цветки мелкие, собраны в короткие, плотные метелки длиной до 10 см. В природе данный вид распространен на Курильских островах, островах Сахалин, Хоккайдо, Хонсю, Корейском полуострове [11, 14]. Вторичный ареал располагается на территории США, Индии, Австралии, Африки и Европы [11]. Для данного вида характерна полиплоидия. Встречаются популяции с наличием тетраплоидного (2n=44), гексаплоидного (2n=66), октоплоидного (2n = 88) и додекаплоидного числа хромосом (2n=132) [10, 11, 15].
В 19 веке данный вид был завезен в Европу в качестве декоративного растения [12]. R. sachalinensis встречается на территории 29 европейских стран, в 18 из них считается инвазивным видом [10, 16]. Данный вид не требователен к составу почв и способен расти практически в любых условиях, образуя монодоминантные заросли [15, 17]. Произрастает по берегам рек, на опушках леса, вблизи дорог, на свалках, строительных площадках, железнодорожных насыпях и заброшенных полях. Ведется поиск мер борьбы с этим растением, а также поиск путей его использования [18].
R. sachalinensis используется как лекарственное, медоносное, декоративное, пищевое и нетрадиционное кормовое растение [19]. Возможно его применение в фиторемедиации загрязненных территорий, в пищевой и бумажной промышленности [20, 21]. Экстракты данного вида используют в качестве стимулятора роста растений и фунгицида [22]. Спиртовое извлечение из листьев оказывает стимулирующее действие при прорастании семян и росте растений пшеницы Triticum aestivum L. [23]. Из экстракта растений R. sachalinensis получен биопестицид [24]. Однако в настоящее время он не был зарегистрирован ни в одной стране ЕС [25]. В Японии и Китае применяется в традиционной медицине [26].
Цель исследования. Данный литературный обзор посвящен систематизации современных научных данных в области химического состава R. sachalinensis, изучению механизмов действия его основных биологически активных соединений и обоснованию их терапевтического использования.
Материалы и методы исследования. Для информационно-аналитического поиска использовали научные данные, размещенные на электронных ресурсах Google Scholar, eLibrary, PubMed, Web of Science, ScienceDirect, Scopus. Поиск осуществляли за период с 2000 г. по 2025 г. по ключевым словам: «Reynoutria sachalinensis», «Fallopia sachalinensis», «giant knotweed», «рейнутрия сахалинская», «ботаническое описание рейнутрии сахалинской», «экстракт рейнутрии сахалинской», «фенольные соединения рейнутрии сахалинской», «химический состав рейнутрии сахалинской», «фармакологическое действие рейнутрии сахалинской», «цитотоксичность», «противовирусное действие», «антибактериальное действие», «противоопухолевое действие».
Профиль биоактивных соединений
Фитохимические исследования рода Reynoutria выявили наличие более 277 химических компонентов [27]. R. sachalinensis содержит большое количество разнообразных биологически активных веществ: фенольные соединения (антрахиноны, стильбены, флавоноиды, фенилпропаноиды, проантоцианидины, гидролизуемые таннины, фенольные кислоты), каротиноиды, хлорофиллы, тритерпеноиды, азотсодержащие соединения и алифатические спирты (табл. 1) [28, 29].
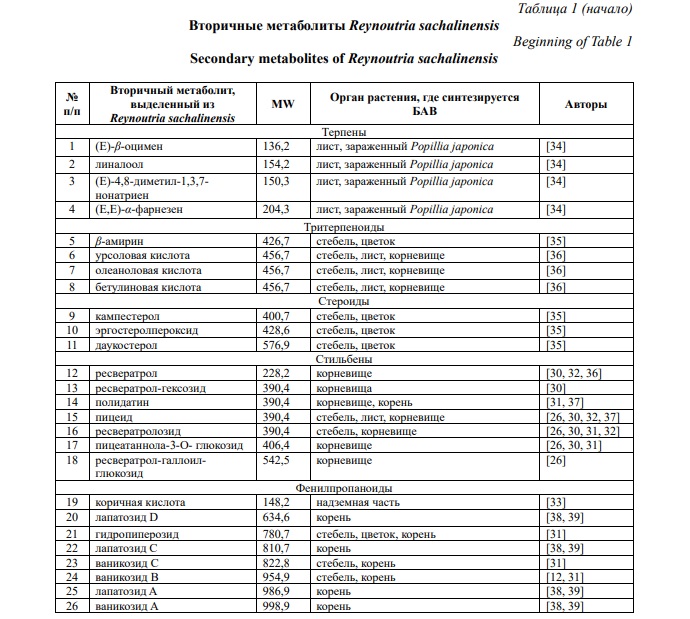
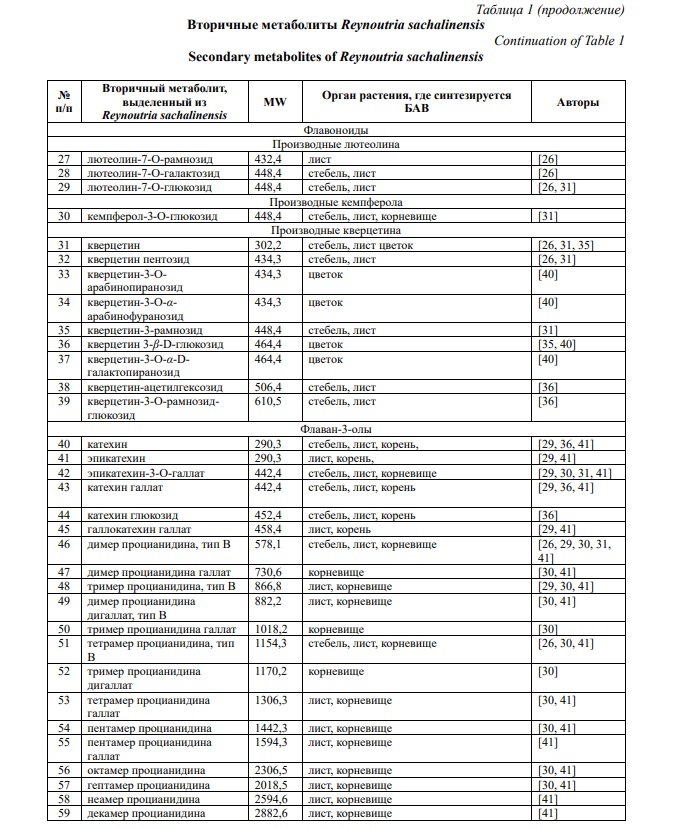
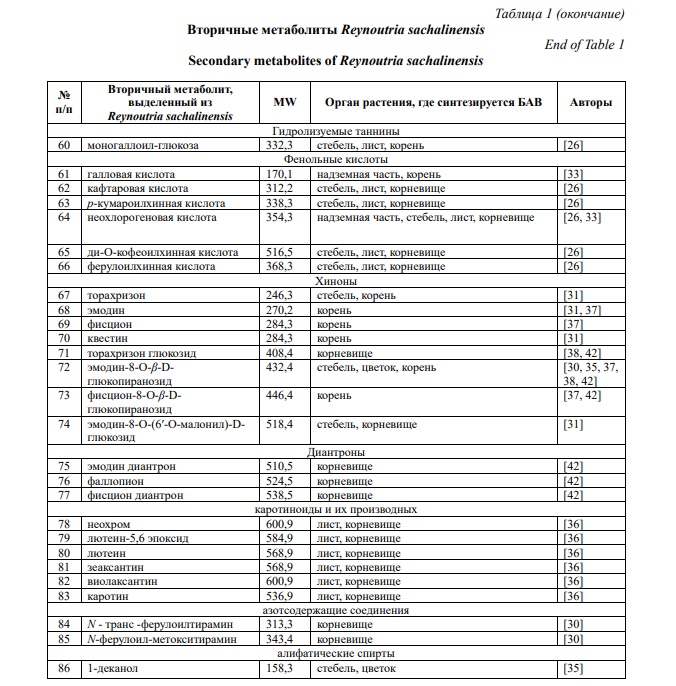
По данным Nawrot-Hadzik I. фракция этилацетата наиболее богата полифенолами (640,7 мг/г) и таннинами (528,3 мг/г) [30]. Содержание стильбенов в корневище R. sachalinensis составляет до 27,04 мг/г, в том числе содержание транс-полидатина – 17,06 мг/г [31]. В молодых весенних побегах накапливается 154 мг/кг ресвератролозида, 64 мг/кг сухой массы пицеида и 68 мг/кг сухой массы ресвератрола [32]. Ваникозид В и С в корнях накапливается до 21,5 и 6,5 мг/г сухого вещества соответственно, а стеблях – 14,3 и 0,6 мг/г сухого вещества [31]. По данным Stafiniak M. с соавторами для R. sachalinensis характерно более высокое содержание гидропиперозида, ваникозидов, чем в R. japonica и R. x bohemica [18]. В надземной части преобладают неохлорогеновая кислота и рутин [33]. Содержание галловой кислоты в надземной части достигает 21,99 мг/кг, что в 3,3 больше по сравнению с корневищем [33]. В листьях R. sachalinensis кверцитрин является основным соединением (до 13,96 мг/г сухого вещества) [31]. По данным Vrchotova N. с соавторами содержание катехина и эпикатехина в молодых весенних побегах достигает 167 и 674 мг/кг сухой массы соответственно [32].
В настоящее время основные исследования направлены на изучение фитохимического состава корневища [29].
Молекулярный докинг вторичных метаболитов R. sachalinensis
Зная пространственную структуру комплексов клеточных белков, мембранных рецепторов и растительных вторичных метаболитов, их лигандов можно предположить механизм их действия, используя молекулярный докинг.
Эфиры фенилпропаноиддисахарида, присутствующие у Reynoutria с преобладающим количеством ваникозидов В и А (рис. 1), до сих пор остаются малоизученными химическими веществами с точки зрения их биологической активности.
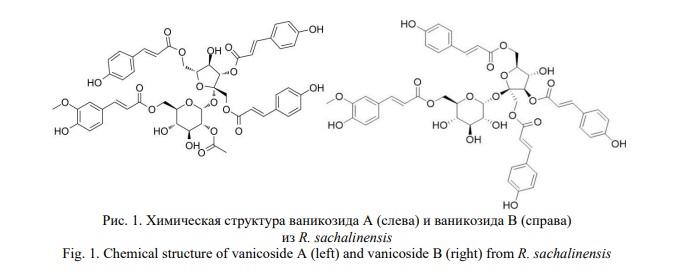
Для оценки возможного противоопухолевого действия проводили молекулярный докинг ваникозидов А и В с активным центром киназы BRAFV600E (ключевой онкоген с мутацией, неправильный сигналлинг BRAF из-за мутации V600E может приводить к избыточной клеточной пролиферации и к ошибочной устойчивости к апоптозу) и MEK-1 (МАР-extracellular regulated kinase) (программное обеспечение GOLD, версия 5.7.2, Кембриджский центр кристаллографических данных, Кембридж, Великобритания) [43] (табл. 2).

Ваникозиды А и В образуют четыре водородные связи с остатками киназы BRAF. Среди них присутствует взаимодействие с остатком Asp594, и это является решающим взаимодействием в случае исходного лиганда. Известно, что аминокислотные остатки Lys483 и Cys532 являются важными для связывания потенциальных ингибиторов BRAF [43].
Ваникозид B взаимодействует с ними обоими, тогда как ваникозид A только с Cys532. В случае МЕК1 оба ваникозида образуют пять водородных связей с этой киназой. Эти результаты позволяют предположить, что оба ваникозида можно рассматривать как потенциальные ингибиторы этих мишеней, а также проводить дальнейшие экспериментальные испытания [44].
Аналогичный эффект наблюдался и в отношении главной протеазы MproSARS-CoV-2 (3CL pro), которая признана ключевой мишенью для профилактики и лечения инфекционных заболеваний, вызванных коронавирусами.
Биологически активные соединения R. sachalinensis были пристыкованы к сайту связывания протеазы SARS-CoV-2 и рассчитано среднеквадратичное отклонение с наилучшим положением и закреплением лиганда (RMSD = 1.6369 Å). В таблице 3 представлены лучшие ингибиторы протеазы (общие взаимодействия с лигандом отмечены жирным шрифтом), балл стыковки больше 90. Известно, чем больше водородных и π-связей, тем устойчивее комплекс [43].
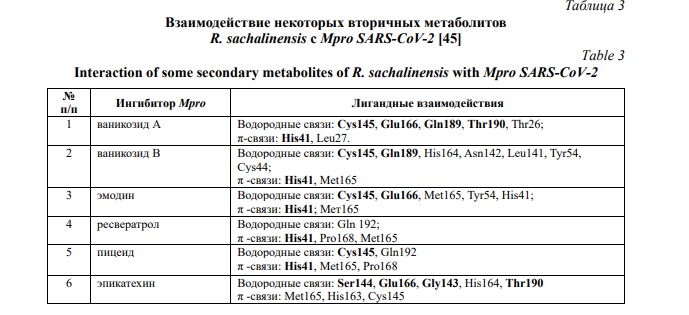
Недавние исследования с использованием молекулярного докинга показали, что стильбены в целом и ресвератрол в том числе, могут быть использованы для лечения COVID-19 [46].
Стильбены, действуя как ингибитор на рецептор ACE2 (ангиотензинпревращающий фермент II), который является функциональным клеточным рецептором для коронавирусов SARS-CoV и SARS-CoV-2 (COVID-19), а также HCoV-NL63 [47, 48, 49], предотвращают образование комплекса S1:ACE2 и проникновение вируса в клетки хозяина.
Исследования молекулярного докинга, проведенные Maroli N. [50], показали, что процианидины также могут быть потенциальными ингибиторами Mpro SARS-CoV-2, а также ACE2 [51, 52, 53, 54, 55].
Биологическая активность экстрактов R. sachalinensis
Спиртовые и водные экстракты R. sachalinensis традиционно используют в странах Восточной Азии для лечения артралгий, кашля, желтухи, аменореи, ожогов, травм, карбункулов и язв [35]. А другой близкий вид семейства Polygonaceae – R. japonica на протяжении многих лет применяют при различных хронических заболеваниях из-за его антиоксидантных, противовоспалительных и противоопухолевых свойств [56]. Вторичные метаболиты R. sachalinensis могут стать источниками для разработки компонентов различных терапевтических препаратов (табл. 4).
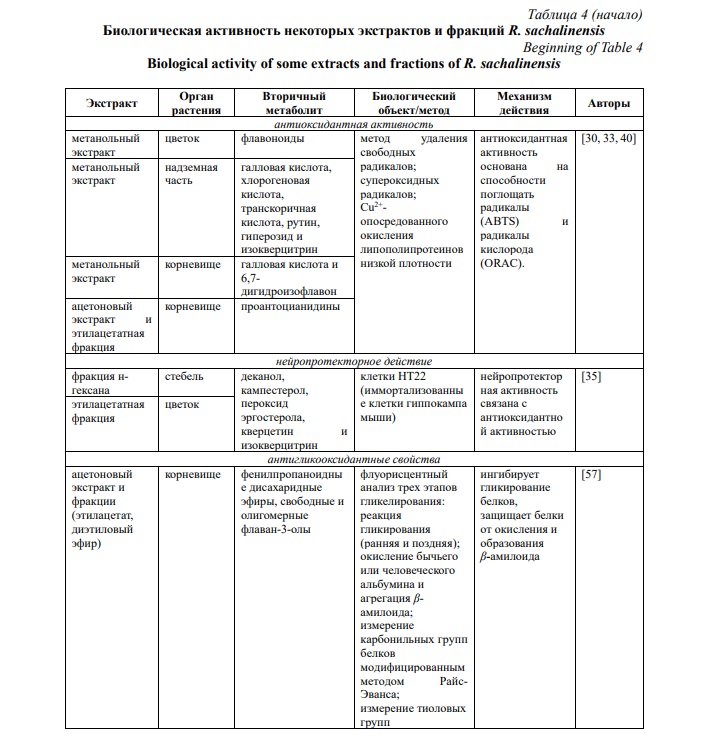
Антиоксидантная активность и нейропротекторное действие. Описана антиоксидантная активность метанольных экстрактов различных органов R. sachalinensis [39]. Исследование антиоксидантной активности методом ВЭЖХ показало, что и надземная часть, и корни имеют антиоксидантный потенциал, который составляет 3,85±0,09 и 3,59±0,09 мг/г в эквиваленте Тролокса соответственно [33]. За антиоксидантную активность отвечают различные биологически активные соединения, содержащиеся в данном виде. Так, выделенные из метанольного экстракта цветков R. sachalinensis флавоноиды кверцетин-3-О-α-L-арабинофуранозид, кверцетин-3-О-β-D-галактопиранозид и кверцетин-3-О-β-D-глюкуронопиранозид показали значительную антиоксидантную активностью со значениями IC50 64,3, 54,7 и 46,2 мкМ (удаление DPPH), значениями IC50 6,0, 6,7 и 4,4 мкМ (удаление супероксидных радикалов) и значения IC50 3,8, 3,2 и 5,4 мкМ против окисления липопротеинов низкой плотности соответственно [40]. Эмодин, фисцион, квестин и их производные также обладают антиоксидантной активностью [31]. Nawrot-Hadzik I. с соавторами показал, что проантоцианидины, представляющие собой фенольные полимеры, в основном состоящие из субъединиц флаван-3-ола, также вносят значительный вклад в общую антиоксидантную способность [30], что может быть обусловлено их восстановительной способностью, связанной с лабильными фенольными гидроксильными группами [61].
В настоящее время профилактика с помощью антиоксидантов является одной из самых инновационных терапевтических стратегий при нейродегенеративных заболеваниях [62]. Eom М. с соавторами также связывают нейропротекторную активность, таких соединений как 1-деканол, кампестерол, пероксид эргостерола, кверцетин и изокверцитрин, выделенных из R. sachalinensis, с антиоксидантной способностью, так как эти соединения снижают уровень активных форм кислорода, а также показали антиоксидантную активность с радикалом DPPH и в анализе поглощения H2O2 [35].
Известно, что ключевую роль в развитии болезни Альцгеймера играет избыточная продукция и накопление β-амилоидного пептида, являющегося основным компонентом сенильных бляшек в ткани мозга. Данный пептид имеет широкий спектр нейротоксических эффектов, включающий окислительный стресс, митохондриальную дисфункцию, нарушение работы ионного транспорта, синаптическую дисфункцию, апоптоз нейронов [63, 64].
В работе Li W. с соавторами показано, что эмодин, который является производным антрахинона и характерен для многих видов семейства Polygonaceae, может быть перспективным кандидатом для лечения болезни Альцгеймера, за счет способности данного соединения препятствовать апоптозу клеток, вызванному β-амилоидом 1-42 in vitro, и β-амилоидом in vivo у трансгенных мышей APP/PS1. Авторы связывают эти эффекты также с антиоксидантной активностью эмодина [49]. Антиоксидантные свойства катехинов, содержащиеся в листьях, стеблях и корнях рейнутрии сахалинской, защищают от нейродегенеративных заболеваний [65] за счет удаления свободных радикалов и регулирования реакции на окислительный стресс [66]. Эпигаллокатехингаллат способствует нейрогенезу гиппокампа взрослого человека, а также играет важную роль в развитии нервной системы и в формировании связей между нейронами [67].
Противовирусная активность. Противовирусная активность R. sachalinensis во многом обусловлена содержанием в различных органах данного вида таких соединений, как эмодин, ресвератрол, ваникозид В и катехинов. Противовирусное действие ресвератрола связано с подавлением внутриклеточного сигнального пути, центральным компонентом которого является транскрипционный фактор NF-κB (nuclear factor κB) [68]. Активность эпигаллокатехин галлата связывают с наличием пирогаллольных и галлоильных фрагментов [69].
Вирус простого герпеса человека. Для эмодина, извлеченного из корневища R. japonica, была обнаружена ингибирующая активность в отношении вируса простого герпеса человека типа 1 у морских свинок [70]. Эпикатехин и эпикатехин-3-галлат также показали высокий уровень ингибирующей активности [71]. Ресвератрол и фитоалексин, относящиеся к стильбенам, могут подавлять репликацию вирусов путем снижения количества вирусных адгезивных клеток и ингибирования реактивации вируса [72]. Установлено, что ресвератрол в отношении вируса простого герпеса типа 1 влияет на экспрессию вирусных генов, синтез ДНК в эпителиальных клетках и регулирует путь серин/треонинкиназа AMP-активируемая протеинкиназа-Sirtuin1 (SIRT1), предотвращая апоптоз в нейронах [73].
Вирус Эпштейна-Барр (гаммагерпесвирус человека 4). Эпигаллокатехин галлат при использовании в концентрациях выше 50 M эффективно подавляет экспрессию литических белков вируса Эпштейна-Барр, но не влияет на экспрессию EBNA-1 [74]. Спиртовой экстракт другого близкого вида (R. japonica) ингибирует литический цикл и снижает выработку вирусных частиц [75].
Вирус Коксаки B4. В работе Liu Z. с соавторами показано, что эмодин, выделенный из R. japonica, в зависимости от концентрации и времени способен ингибировать проникновение и репликацию вируса Коксаки B4 [76].
Короновирусы. Как известно инфекция SARS-CoV-2 начинается с проникновения вируса в клетку посредством взаимодействия вирусного белка Spike (S) с рецептором клеточной поверхности ACE2 (ангиотензинпревращающий фермент 2) и праймирования белка S клеточной протеазой TMPRSS2 (трансмембранная сериновая протеаза 2) [77]. Протеолитическая обработка белка S TMPRSS2 приводит к образованию фрагментов S1 и S2, которые остаются нековалентно связанными. Полипептид S1 взаимодействует с рецептором ACE2 через домен связывания рецептора (RBD), а полипептид S2 облегчает слияние вируса с клеточной мембраной [78]. Интернализованный вирус транслирует РНК-геном в полипептид, за которым следует высокорегулируемая и скоординированная протеолитическая обработка основной протеазой (Mpro/3CLpro), химотрипсин-подобной протеазой, и расщепление субстратных полипептидов [79, 80]. Блокирование процесса созревания вирусных полипротеинов нарушает репликацию вируса в клетках-хозяина [81], поэтому вирусные и клеточные белки представляют собой мишени, воздействуя на которые становится возможным лечение и профилактика SARS-CoV-2.
Установлено значительное ингибирование Mpro SARS-CoV-2 при действии ацетонового экстракта R. sachalinensis, характеризующегося высоким содержанием процианидинов и фенилпропаноидных эфиров дисахарида [45, 51, 82, 83].
Бутанольные фракции R. sachalinensis показали ингибирование Mpro SARS-CoV-2 с IC50 = 4,031 мкг/мл [45]. В работе Yang M. с соавторами показано, что ресвератрол способен значительно ингибировать репликацию SARS-CoV-2 с EC50 = 4,48 мкМ [84]. Водные и этанольные экстракты R. japonicaзначительно ингибируют проникновение псевдовируса SARS-CoV-2 за счет блокирования взаимодействия белка ACE2/S и ингибирования протеазы 3CL [85].
Эмодин способен ингибировать взаимодействие спайкового белка SARS-CoV и человеческого ACE2 [52] и нарушать выход вируса SARS-CoV и HCoV-OC43, благодаря блокировке ионного канала, посредством которого происходит высвобождение вируса из инфицированной клетки [86].
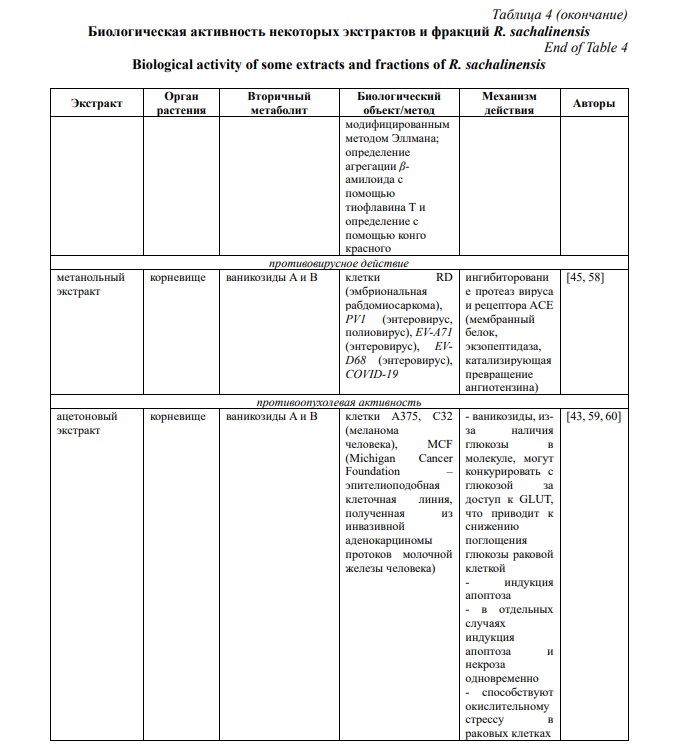
Эпигаллокатехин галлат отвечает за подавление активности главной протеазы в SARS-CoV-2 и репликации вируса in vitro [87]. Ваникозид В также оказывает умеренное ингибирующее действие на протеазы вируса SARS-CoV-2 [45].
Установлено, что олигомерные проантоцианидины обладают ингибирующей активностью в отношении протеиназы TMPRSS2 [88].
Вирус иммунодефицита человека. Ресвератрол и эмодин-8-O-β-D-глюкозид способны ингибировать образование синцития, вызванное ВИЧ-1 с EC50 = 4,37 и 11,29 мкг/мл соответственно [89, 90]. Эпигаллокатехин галлат является ингибитором обратной транскриптазы ВИЧ, ингибирует проникновение ВИЧ в клетки-мишени и вызывает снижение способности вирионов инфицировать клетки [91].
Вирус гепатита C. В работе Calland N. с соавторами показано, что (2)-эпигаллокатехин-3-галлат, эпикатехин-3-галлат и эпигаллокатехин обладают противовирусной активностью HCV на ранней стадии жизненного цикла вируса [92]. Установлено, что (-)-эпигаллокатехин-3-галлат подавляет вирус, прикрепляясь к целевой клетке и предотвращая распространение инфекции на другие клетки [92].
Энтеровирус. Эпигаллокатехин галлат и галлокатехин галлат способны подавлять репликацию энтеровируса EV71. Установлено, что ваникозид B защищает клетки RD (эмбриональная рабдомиосаркома) от EV-A71 и EV-D68 при концентрации 20 мкМ [45], а также является ингибитором активности фермента, необходимого для репликации EV.
Полиовирус. В работе Arita М. указывается, что ваникозид В, выделенный из метанольного экстракта корня R. sachalinensis, обладает антиполиовирусным действием [46, 93], ингибируя фермент PI4KB (фосфатидилинозит-4-киназа бета), который является его прямой мишенью [93]. Однако антипролиферативный эффект на лимфоциты [94] и летальность у мышей [95] вызывают сомнения по поводу безопасности использования соединния in vivo.
Противоопухолевая активность и цитотоксическое действие. Известно действие ваникозидов А и В, выделенных из корневищ Reynoutria sachalinensis, на две линии клеток меланомы человека (амеланотическая – C32 и меланотическая – A375, которые содержали эндогенную мутацию BRAFV600E) и две нормальные клеточные линии человека (кератиноциты (HaCaT) и фибробласты). Ваникозид А (5,0 мкМ) показал более сильную цитотоксичность в отношении клеточной линии C32, при этом жизнеспособность клеток снизилась на 45% уже после 72-часовой инкубации, что возможно объясняется дополнительной ацетильной группой в молекуле [43].
Механизм действия ваникозидов на клетки меланомы более сложный, чем просто индукция апоптоза [60]. В некоторых случаях, таких как повреждение клеток кислородными радикалами, два механизма гибели клеток – апоптоз и некроз – происходят одновременно. Кроме того, ваникозиды, как и другие полифенолы, могут способствовать окислительному стрессу в раковых клетках [60].
Ваникозиды вызывают гибель клеток меланомы в концентрациях от 2,5 до 50 мкМ, не нанося вреда первичной линии фибробластов. Линия клеток кератиноцитов HaCaT более чувствительна к ваникозидам, чем фибробласты, показывая явное снижение жизнеспособности после инкубации с 25 мкМ ваникозида А, но без измеримой флуоресценции, связанной с гибелью клеток. Поэтому необходимы дальнейшие исследования, чтобы гарантировать селективность в отношении раковых клеток и безопасность [43].
В настоящее время известно, что ваникозиды проявляют цитотоксичность и на клеточную линию MCF (Michigan Cancer Foundation – эпителиоподобная клеточная линия, полученная из инвазивной аденокарциномы протоков молочной железы человека) при субмикромолярных дозах [96]. А Ваникозид B ингибирует двухэтапный канцерогенез опухоли кожи у мышей, индуцированной 12-O-тетрадеканоилфорбол-13-ацетатом, путем подавления активности фермента циклин-зависимой киназы [97].
Прочие виды активности. Фенилпропаноидные гликозиды ваникозидов A и B обладают ингибирующим действием на β-глюкозидазу [40, 58], метанольный экстракт и изокверцитрин ингибируют активность ацетилхолинэстеразы и α/β-глюкозидазы [35, 39], метанольные экстракты корней и листьев – активность ксантиноксидазы [98], 3-О-β-D-галактопиранозид кверцетина, лапатозид D и N-транс-ферулоилтирамин – активность акарбозы [39], торахризон и торахризонглюкозид способны ингибировать α-амилазу [31]. Эмодин, фисцион, квестин и их производные обладают противомикробной и противовоспалительной активностью [31].
Таким образом, растительное сырье вида R. sachalinensis можно рассматривать как источник биологически активных метаболитов, имеющих подтвержденный широкий спектр использования, в том числе противовирусное и противоопухолевое средство, но необходимы дальнейшие работы по выделению и наработке значимых вторичных метаболитов и оценке их механизма действия invitro и invivo.
Информация о финансировании
Работа выполнена согласно государственному заданию в ФГБНУ ВИЛАР «Исследование механизма синтеза конденсированных таннинов лекарственных растений и их биоактивные свойства FGUU-2025-0005».














Список литературы