
A replication study of genetic variants associated with high-level musical aptitude
Abstract
Background: Existing genetic studies on musical abilities have focused on various molecular approaches, which determined gene regions/variants implicated in (i) music perception and performance based on genome-wide linkage and association scans, (ii) differential expression patterns as a response to music listening/performance, (iii) music production in songbirds, and (iiii) neuronal plasticity relation to music training. The aim of the study: The present study sought to examine the main and combined effects of related gene variants in developing musical abilities in individuals from Russia. In addition, we aimed to obtain the mathematical model with the optimal sensitivity to specificity ratio to predict individual liability for high-level musical aptitude. Materials and methods: A genotyping of the SNCA (rs356168, rs3910105), GATA2 (rs9854612), PCDH7 (rs13109270, rs13146789), UNC5C (rs9307160), ASAP1 (rs3057), SBSPON (rs1007750), GRIN2B (rs3764030), and BDNF (rs6265) gene variants was performed via real-time PCR with endpoint fluorescent detection in high-level musically skilled students (N = 100, 66% of women) in Moscow (Russia) and corresponding control group (N = 200, 67% of women). The main and combined effects of examined gene variants on developing musical abilities were assessed via logistic regression models followed by a selection of the best predicting model in R v.4.1.2. Results: We obtained a regression model (AUC = 0.735, P = 0.026), which can predict a higher probability to develop musical aptitude linked to SNCA rs356168 TT-genotype (β=1.98, P=0.011, OR=7.31, 95%CI 1.66-38.4), SNCA rs3910105 TC-genotype (β=1.39, P=0.034, OR=4.16, 95%CI 1.2-16.6), GATA2 rs9854612 A-allele (β=0.83, P=0.082, OR=2.30, 95%CI 0.91-6.11). Moreover, a significant effect of ASAP1 rs3057 on manifestation of musical skills was observed under the dominance deviation from additivity model (β=2.13, P=0.032, OR=1.94, 95%CI 1.05-3.56). Conclusion: Revealed findings testify in the involvement of dopaminergic neurotransmission and reward mechanisms attributed to α-synuclein- and GATA2-mediated gene effects in ability to develop exaggerated musical skills.
Keywords: musical abilities, absolute pitch, synuclein, neurotransmission, brain functioning, regression model, ROC-analysis
Background. Musical perception/performance is a complex cognitive skill representing a unique feature of humankind. The musical aptitude, which includes the ability to perceive and understand intensity, timbre and tone duration, pitch, rhythm and structure [1], existed even ~ 40,000 years ago based on the archaeological excavations of old flutes dated to this time [2]. It has been suggested that musical aptitude has been evolved due to its favorable effect on negative emotional states, from the one side, and to the activation of brain reward circuitry like the limbic and mesolimbic structures, from the other side [3]. Music performance relies on a sensory perception, execution of cognitive tasks, emotions, memory and learning [3], thus indicating a complicated interaction of various neurobiological systems.
According to neurobiological hypothesis, the development of musical abilities as a cognitive-related trait [4] is a complex process affected by multiple genes, epigenetic and environmental components. Twin research demonstrated that heritability estimates accounted for 0.21-0.68 depending on certain musical aptitudes [5]. Together with different types of personality abilities attributed to brain functioning such as mathematical abilities [6], spatial abilities [7], personality characteristics [8, 9], musical aptitude represents a complex phenotype caused by the interaction of multiple genes and environmental factors [10].
During recent decades the study of a genetic basis for developing musical skills has become a subject of growing curiosity. Initially, the role of hereditary component has been established while examining a large pedigree of musically talented individuals from Finnish ancestry [11]. Subsequently, genome-wide linkage analyses determined a significant evidence for linkage for such genomic loci as 4q22 (at rs9307160 in the UNC5C gene) for music perception [11, 12], 8q24.21 (at rs3057 in the ASAP1 gene), 8q21.11 (at rs1007750 in the SBSPON gene) for absolute pitch [13]. Genome-wide association studies (GWAS) revealed the association of rs9854612 (GATA2, 3q21.3,), rs13146789 and rs13109270 (PCDH7, 4p15.1) with music perception [5]; whereas transcriptome-based analyses demonstrated a differential expression of α-synuclein gene (SNCA, 4q22) while listening to classical music [10, 14, 15, 16]. Some studies implicated neuronal plasticity genes, for instance, brain-derived neurotrophic factor (BDNF) [17] and GRIN2B genes [1] to be related to music ability.
In summary, published findings, which depicted gene regions/variants implicated in (i) music perception and performance based on genome-wide linkage and association scans, (ii) differential expression patterns as a response to music listening/performance, (iii) music production in songbirds, and (iiii) neuronal plasticity in relation to music training, may represent the basis for the analysis of these gene variants in manifestation of musical aptitude in humans.
The aim of the study. Considering scarce data on the replication of genetic effects on developing musical aptitude obtained via different methodological approaches, in the present study we sought to examine the main and combined effects of related gene variants in a higher probability to develop musical abilities in individuals from Russia. In addition, we aimed to obtain the mathematical model with the optimal sensitivity to specificity ratio to predict individual liability for high-level musical aptitude.
Materials and methods. The study sample included musically talented students, who underwent a severe contest including pitch-production accuracy test to pass the examination for the study at the musical conservatories (N = 100, 66% of women; aged 18-22 years) in Moscow (Russia). All enrolled individuals were ethnically Russians. A control group included mentally healthy individuals (N = 200, 67% of women; aged 18-23 years) corresponding by age, sex, and ethnicity to the group of musically talented students. All the enrolled individuals reported no family history of mental disorders and had not been recorded in the Psychiatric Registry. The participants for the study signed an informed consent, which stated a voluntary principle for the participation in the scientific research and reported a detailed explanation of all the procedures. The study was approved by the local bioethical committee at the Institute of Biochemistry and Genetics (Ufa, Russia) in accordance with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
DNA extraction via phenol-chloroform technique was followed by genotyping using PCR with end-point KASP fluorescent detection (MaxMedical, Moscow, Russia) on CFX96 DNA-Analyzer (BioRad, USA). A selection of SNPs was based on previous findings, which indicated the association or linkage of SNPs in musical aptitude or related phenotypes such as language and absolute pitch. A complete list of SNPs included those located in the SNCA (rs356168, rs3910105), GATA2 (rs9854612), PCDH7 (rs13109270, rs13146789), UNC5C (rs9307160), ASAP1 (rs3057), SBSPON (rs1007750), GRIN2B (rs3764030), and BDNF (rs6265) genes. The data on their chromosomal position, minor allele frequencies, and the Hardy-Weinberg equilibrium (HWE) test are shown in Table 1. Allele and genotype frequencies distributions of all the examined gene variants were congruent with the HWE test except for PCDH7 rs13109270 (P < 0.001), which has been excluded from all the subsequent statistical procedures.
After the testing for the correspondence of the distribution of allele and genotype frequencies to the HWE, statistical analysis has been carried out, which included the estimate of the main effect of gene variants on individual liability to develop high-level musical aptitude in the total sample with sex and age inclusion as covariates. The main effects were assessed via logistic regression under additive, dominant, recessive, and dominance deviance from the additivity (domdev) models with control for sex and age (PLINK v.1.09). In order to detect the possible haplotypes association in the SNCA gene, a linkage disequilibrium examination (based on D’ and r2) followed by haplotypes association analysis was performed (PLINK v.1.09). To estimate the effect of multiple gene variants on developing musical abilities we designed logistic regression models (under general linear models command) with inclusion of all examined SNPs followed by a selection of the best predicting model based on the procedure of backward elimination, which was chosen on the basis of the best Akaike information criterion (AIC) and p-value in R (v.4.1.2). In the case of low frequency of a minor homozygote in all examined loci, we performed the analysis between minor allele (grouped minor allele homozygote and heterozygote) and major allele homozygote genotypes. The ROC-curve (Receiver Operating Characteristic) construction and AUC (area under curve) calculation were performed in R (v.4.1.2).
Results and discussion. In the present study we sought to untangle the main effect of each selected gene variant on developing high-level musical aptitude. Based on the results of logistic regression, which was conducted with sex and age inclusion as covariates and analyzed different models of allele effects, a significant effect of ASAP1 rs3057 on manifestation of specific musical abilities was observed under the dominance deviation from additivity model (β = 2.13, P = 0.032) (Table 1). To be more precise, the frequency of heterozygous ASAP1 CT genotype (OR = 1.94, 95%CI 1.05-3.56) was higher in musically talented students compared to CC and TT homozygotes. Other SNPs failed to significantly affect liability to musical aptitude. However, there was a trend toward an increased frequency of rs356168 major TT genotype in the SNCA gene (β = 1.80, P = 0.071, OR = 1.49, 95%CI 0.96-2.30) in music-sensitive individuals compared to control group, whereas rs356168 AA genotype demonstrated a trend to be a marker of liability to musical abilities (Table 1).
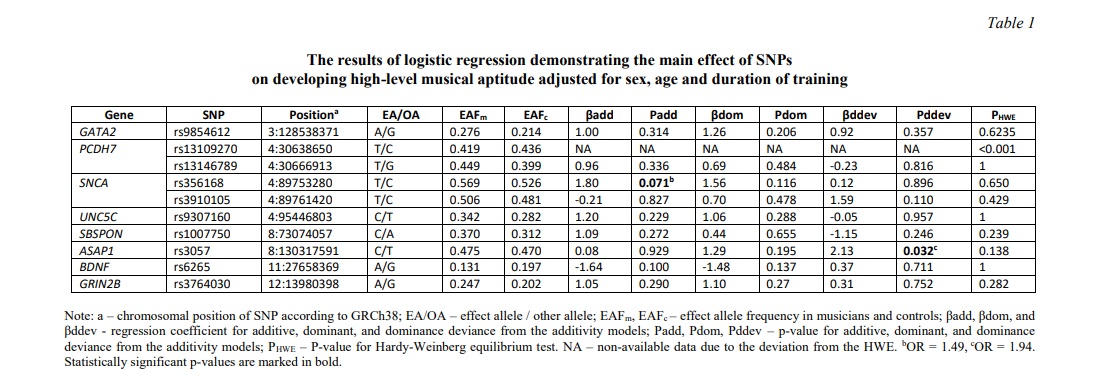
Since we included genetic loci located in the same genes (i.e. rs356168, rs3910105 in the SNCA gene, and rs13109270, rs13146789 in the PCDH7 gene), we aimed to estimate the possible haplotype association with musical abilities. However, due to the deviation of PCDH7 rs13109270 from the HWE, we carried out linkage disequilibrium analysis in the SNCA gene only. The rs356168 and rs3910105 have been moderately linked (r2 = 0.20, D’ = 0.49) and were located at a distance of ~ 8,140 bp. At a subsequent stage we calculated haplotypes frequencies in the SNCA gene (on the basis of rs356168 and rs3910105), which corresponded to 0.372 (TC-haplotype), 0.347 (CT-haplotype), 0.166 (TT-haplotype), and 0.115 (CC-haplotype). As a result of haplotypic analysis we revealed a trend for association of SNCA TT-haplotype with enhanced musical abilities (β = 3.49, P = 0.061, OR = 1.85). However, this association failed to achieve the level of statistical significance.
In order to examine a combined effect of various gene variants on liability to develop high-level musical skills and to construct mathematical model with high sensitivity to specificity ratio to predict developing musical aptitude, we carried out a series of logistic regressions (under general linear models procedure). At the initial stage all the SNPs, as well as sex and age have been included as potential covariates. However, a statistically significant effect was demonstrated only for GATA2 rs9854612 (β = 1.18, P = 0.038) and SNCA rs356168 (β = 2.26, P = 0.012), while other predictors appeared to be non-significant, thus resulting in a non-significant value of the model itself (P = 0.059) (Model 1, Table 2). Subsequently, as a result of backward elimination function we obtained the final model (Model 2, Table 2), which may significantly predict individual’s liability to develop high-level musical skills (P = 0.026). According to this model, a higher probability to develop musical aptitude was associated with SNCA rs356168 TT-genotype compared to CC-genotype (β = 1.98, P = 0.011, OR = 7.31, 95%CI 1.66-38.4), SNCA rs3910105 TC-genotype compared to TT-genotype (β = 1.39, P = 0.034, OR = 4.16, 95%CI 1.2-16.6), GATA2 rs9854612 A-allele (β = 0.83, P = 0.082, OR = 2.30, 95%CI 0.91-6.11) compared to GG-genotype. Although other genotypes of the SNCA rs3910105 and rs356168 demonstrate a non-significant effect (P < 0.05), they have to be controlled for in the regression models. In order to estimate a prognostic value of the obtained model, we constructed a ROC-curve with a moderate measure of classifier performance (AUC = 0.735) (Fig. 1).


Assuming that musical aptitude is a complex trait, which is manifested as a result of genetic interactions of small effect and specific predisposing environment, we examined a simultaneous effect of multiple gene variants in a summarized model. In total, regression model obtained by our research group evidence toward a combined effect of SNCA rs3910105 and rs356168 and GATA2 rs9854612 on a liability to develop high-level musical aptitude. Previous research indicated exaggerating expression of the SNCA gene as a response to music listening [14, 15, 16]. Namely, transcriptome studies indicated overexpression of 45 genes, including the SNCA gene, as a result of 20 min listening to Mozart Violin Concerto No 3 independently on the participant’s experience [15]. Subsequent study of mentioned research group verified an exaggerated expression of the SNCA gene caused by 2h listening to classical music in professional musicians compared to control group [16]. At the same time, a decreased abundance of SNCA transcripts were characteristic for neurodegenerative states such as Parkinson’s disease [18]. Since synucleins, which are responsible for the regulation of synaptic vesicle trafficking and neurotransmitter release, are abandoned on presynaptic dopaminergic neurons and mediate dopaminergic neurotransmission [18], our data also confirm the role of music performance in dopaminergic reward mechanisms. The SNCA rs3910105 and rs356168 examined in the present study have been demonstrated to affect expression of SNCA transcripts (cis effect) and several other genes in trans in individuals with Parkinson’s disease (PD) [19]. Namely, SNCA-007 transcript upregulation in PD was related to rs356181 CC, rs3910105 CT and TT genotypes, while elevated levels of SNCA-008 transcript were characteristic of rs356181 CT and TT variants together with rs3910105 GG genotype [19]. Therefore, a genetic effect of rs356181 and rs3910105 genotypes on transcription of various SNCA isoforms in PD appears to be in opposite direction. It should be noted that mentioned transcripts SNCA-007 and SNCA-008 represent truncated isoforms of SNCA mRNA (skipping one or several exons) prevalent in neurodegenerative states and represent biomarkers of Parkinson’s disease [18, 20]. To date no studies reporting a link between SNCA expression and rs3910105 / rs356168 allelic variants outside synucleinopathies was published, therefore, we cannot make unambiguous conclusion on a direction of such association. Nevertheless, our findings indicate that higher ability to develop musical skills was linked to SNCA rs356168 TT and SNCA rs3910105 TC genotype under a multiple regression model.
The α-synuclein is involved in regulation of dopamine biosynthesis and terminates dopamine neurotransmission via alteration of dopamine transporter (DAT)-mediated dopamine uptake from the synapses [21]. In this regard interesting findings indicated a link between DAT availability and SNCA gene variants in healthy subjects [20]. To be more precise, a trend for diminished DAT availability in striatum was shown for rs3910105 TT genotype carriers compared to individuals with CC/TC genotypes. These results seem to be in line with our data, which detected higher musical skills to be associated with rs3910105 TC compared to TT genotype, since better musical performance was hypothesized to be linked to enhanced dopaminergic neurotransmission. Another participant of dopaminergic signaling such as GATA2 protein was shown to regulate the expression of the SNCA gene and is highly abandoned in dopaminergic neurons [14], and was also implicated at a trend level in musical skills development in the present study. A functional role of GATA2 in sounds recognition and music perception has been previously shown: GATA2 is involved in tonopic mapping due to its role in the development of cochlear hair cells and inferior colliculus [22]. Although other participants of dopaminergic signaling may play a role in SNCA-mediated impact on developing musical abilities, since dopaminergic functioning is also attributed to genetically-caused variance in DRD2 and SLC6A3 gene expression [23], we failed to detect the association of other participants of dopaminergic signaling (i.e. BDNF, GRIN2B genes) with the examined phenotype.
Another finding of the present study is the association of a heterozygous ASAP1 rs3057 CT genotype with musical aptitude. The ASAP1 gene encodes Arf GTPase-activating protein (GAP), which mediates neuronal plasticity and regulates membrane trafficking and cytoskeleton remodeling [24]. The rs3057, which has previously demonstrated a linkage with absolute pitch (exponential LOD score = 3.464, empirical genome-wide p = 0.03) in Europeans [13], confirmed the association with musical aptitude in the Russian cohort. However, rs3057 has been reported to neighbor the ADCY8 gene encoding adenylate cyclase 8 [13], which is abundantly expressed in the brain and is implicated in cognitive processes [25], a real distance between rs3057 and the ADCY8 gene is more than 462 kb. Actually, this SNP resides an intronic region of the ASAP1 gene and is located in a close proximity to several microRNA-encoding genes. However, the functional role of rs3057 remains unknown. Nevertheless, the involvement of ASAP1 gene in cognitive-related traits including musical abilities seems to be plausible, since the ASAP1 gene is highly expressed in the brain. Moreover, the involvement of ASAP1 gene in some pharmacological issues of therapy of such mental-related phenotype as ADHD has been indicated [26].
Other examined gene variants, including PCDH7 rs13109270, rs13146789, UNC5C rs9307160, SBSPON rs1007750, GRIN2B rs3764030, and BDNF rs6265 failed to demonstrate their association with musical skills in the analyzed cohort of professional musicians. However, previously, variations in protocadherin gene cluster 199 (PCDHA1-9), which is involved in serotonergic functioning in the brain and synaptogenesis, were related to music ability [27]. Despite the initial report of UNC5C rs9307160 and SBSPON rs1007750 linkage with music perception and absolute pitch [11, 12, 13], no subsequent studies confirmed the involvement of these SNPs in the development of music- or cognitive-related phenotypes. In addition, the studies unraveling the genetic basis of song perception and production in songbirds (zebra finch) succeeded to detect the impact of GRIN2B gene [1], which encodes a subunit of N-methyl-D-aspartate (NMDA) receptor (subtype of ionotropic glutamate-gated ion channel) involved in synaptic plasticity and learning. Although a recent study reported the role of GRIN2B rs3764030 located in the promoter gene region and GRIN2B A-allele, which is associated with elevated GRIN2B mRNA levels, in mental processing speed [28], the effect of GRIN2B gene variants on developing musical skills appeared to be non-significant in the present study, as well as in our previous study examining genetic role in spatial abilities [7].
Some studies implicated neuronal plasticity genes, for instance, brain-derived neurotrophic factor (BDNF) [17] to be related to music ability. Musical talent can be partially attributed to the efficacy of innate ability for motor and sensory learning to play musical instruments. From the other side, highly effective musical performance is related to both early onset of music training and increased expression of neuronal plasticity genes, for instance, BDNF. Animal studies also evidence in the positive effect of music listening on a diminished anxiety and BDNF overexpression even in BDNF Met/Met knock-in transgenic mice [29]. In turn, a recent pilot study [17] suggested the presence of a compensatory effect of early music training, which promoted enhanced sensorimotor learning in individuals with BDNF Met-allele. In line with this observation, repetitive training had a positive effect on cognitive functioning only among Met-allele carriers of younger age [30]. Therefore, it seems that a positive effect of music training is essential for individuals with BDNF-linked deficit of neuronal plasticity (i.e. Met-allele). Although in the present study minor allele frequency of BDNF Met-allele was lower among musicians (0.131) compared to control group (0.197), this difference was statistically non-significant and, therefore, a beneficial impact of training in Met-allele carriers was not controlled for.
Conclusion. In conclusion, this preliminary study, which aimed to estimate the presence of genetically-mediated cause of developing high-level musical skills in a relatively moderate but strongly selected sample of professional musicians, revealed a combined effect of SNCA (rs3910105, rs356168) and GATA2 rs9854612 gene variants on liability to develop high-level music skills, which was based on the established significant regression model characterized by a moderate measure of classifier performance. The model predictors stressed the role of SNCA gene variants associated with exaggerated dopaminergic neurotransmission in developing musical talent. Moreover, we succeeded to demonstrate the impact of ASAP1 rs3057 in musical abilities; however, the studies unraveling the functional link between rs3057 allelic variants and changes in gene expression in cis or in trans have to clarify detected association.
Financial support
The study was supported by the State Contract of the Ministry of Science and Education of RF (№АААА-А16-116020350032-1) with partial support of the Russian Science Foundation (project no. 17-78-30028) (in the part of collection of biological materials) and the Ministry of Science and Higher Education of the Republic of Bashkortostan (agreement no. 1, December 28, 2021) (in the part of molecular-genetic analysis). DNA samples for the study were provided by the IBG UFRC RAS collection “Collection of human biological materials” developed within the project № 007-030164/2.
Thanks
The study was supported by the State Contract of the Ministry of Science and Education of RF (№АААА-А16-116020350032-1) with partial support of the Russian Science Foundation (project no. 17-78-30028) (in the part of collection of biological materials) and the Ministry of Science and Higher Education of the Republic of Bashkortostan (agreement no. 1, December 28, 2021) (in the part of molecular-genetic analysis). DNA samples for the study were provided by the IBG UFRC RAS collection “Collection of human biological materials” developed within the project № 007-030164/2.














Reference lists